|
Spermathèque des Telemidae femelles
ANATOMIE ET COMPORTEMENT DES ARAIGNEES :VINGT-CINQ
ANS DE RECHERCHES
Par André LOPEZ, auteur (version 2024) |
|
Couleurs
conventionnelles :
Couleurs
conventionnelles : En noir et italiques, termes anatomiques ; en violet,,
noms
génériques et spécifiques ; en vert, noms de
familles et sous-familles ; en
orange,, parties
les plus importantes
et résumés; en bleu, liens divers.
|
|
Abréviations
conventionnelles :
M.E.B. : (photographie en) microscopie électronique à balayage M.E.T. :
(photographie en) microscopie
électronique à transmission
C.H. : coupe histologique
(microscopie photonique)
|
1 - Introduction
2 - Structure histologique
3 - Ultrastructure
3-1- Paroi de la
spermathèque-
3.1.1 -
Coussinet
3.1.1.a -
Epithélium
3.1.1.b -
Cuticule du coussinet
3.1.2 - Corps
de la spermathèque
3.1.2.b -Cuticule
3.1.2.c - Espace sous-cuticulaire
3.1.2.d - Musculeuse
4 - Commentaires
1-Introduction
On sait que l'orifice génital primaire ou gonopore des Araignées femelles s’ouvre, comme celui des mâles, au milieu de la fente ou sillon épigastrique marquant la limite entre les 2eme et 3eme somites opisthosomiens (abdominaux).
Très
simple
chez les seconds, où
il
ne présente que les différenciations superficielles de l'appareil épigastrique, il est
également unique et banal chez les femelles de
certains groupes “primitifs”, Orthognathes et Araignées dites
"Haplogynes"dont font partie les Telemidae, en
particulier Telema tenella,
leur “chef de file”. L’orifice
génital n’y est
pas surmonté par
une épigyne.
Celui
de Telema s’ouvre dans l’atrium génital,
sillon épigastrique devenu
largement
béant. Il
lui fait suite un uterus
externus impair et médian recevant deux oviductes qui prolongent
eux-mêmes les
ovaires. A l’atrium est annexée
une unique
spermathèque (réservoir
spermatique).
Il s’agit d’un sac impair, allongé (Fig.1) et «extraordinairement volumineux » suivant l’expression de Simon et Fage (1922) qui l’ont décrit pour la première fois dans des préparations éclaircies.
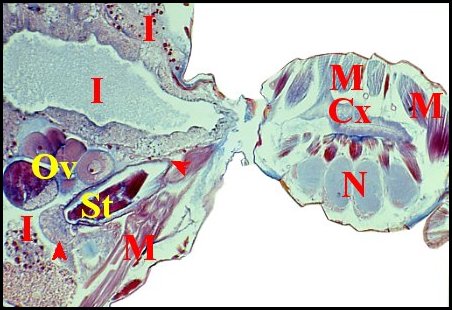 |
| Fig.1-
Telema
tenella, femelle : coupe longitudinale, un peu oblique pour
le prosoma (à droite) |
| Cx, glande coxale (prosoma) - I, intestin et diverticules -M, muscles - N, ganglions nerveux - Ov, ovaire - St, spermathèque à extrêmités fléchées.(© A.Lopez C.H.). |
Avant que l'auteur ne publie deux photos
sommaires de la spermathèque chez
Telema tenella (Lopez,1976
: p.26, fig.6 et 7) en signalant qu’elle loge
des spermatophores, il n’existait aucune représentation
histologique de cet organe dont
les figures étaient purement macroscopiques et à peine esquissées (Fage,1913 ; Simon & Fage,1922).
L’année suivante, Lopez, (1977) décrivit pour la première fois sa structure en détail et, plus tard, son aspect électronomicroscopique en collaboration avec L.Juberthie-Jupeau (Lopez,1983b)
Les deux
chercheurs ont
utilisé comme matériel d'études l’espèce Telema tenella Simon (Telemidae)
récoltée (3 eme photo)
dans
une grotte des Pyrénées orientales (Can Britxot, près de La Preste), préparée pour l’histologie
selon des techniques banales et
transportée également vivante au Laboratoire ariégeois de Moulis (Note 1) pour l’étude ultrastructurale.
La
spermathèque de
Telema est un
organe intra-abdominal
médian, allongé selon le grand
axe de l’opisthosoma,
surmonté par les ovaires et
les diverticules intestinaux,
reposant sur
les glandes à
soie les plus
antérieures
(ampullacées
notamment) qu’elle
refoule. Subcylindrique et sacciforme, elle mesure
environ 300 µm
de long et 80 µm de diamètre. Son extrémité antérieure s’ouvre dans l’atrium génital et
son extrémité
postérieure se
termine en un cul de sac plus ou moins incurvé.
Sa cavité, de section plus ou moins arrondie, renferme, chez les femelles fécondées, de la sécrétion et un spermatophore qui s’y love en se repliant plusieurs fois sur lui-même.
Sa
paroi comporte plusieurs
couches : une couche
musculaire externe striée,
un épithélium
simple et une cuticule interne continue.
Au niveau du corps de la spermathèque, cette cuticule a une épaisseur moyenne de 3 µm ; l’épithélium est assez clair, pourvu de noyaux arrondis, haut de 14 µm sur la face ventrale et de 5 µm sur les faces dorsale et latérales.
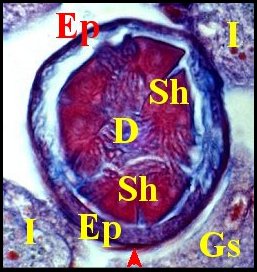 |
 |
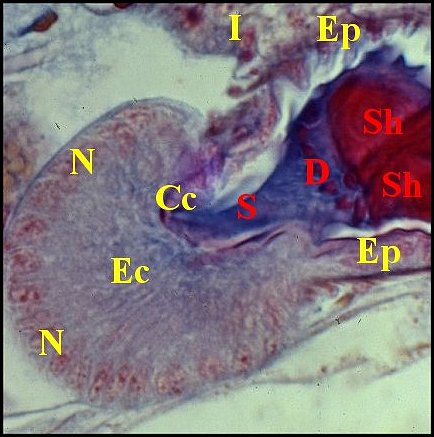 |
| Fig.2- Telema : spermathèque, coupe transversale | Fig.3- Telema : spermathèque,
coupe longitudinale partielle |
Fig.4- Détail :
coussinet |
| Cc, cuticule -
Ct, coussinet - D, digitations des spermatozoïdes - Ec, épithélium du
coussinet - Ep, épithélium du corps de la spermathèque - Gs, glande à soie - I, intestin
-L, lumière de la spermathèque - M, muscle hors
spermathèque -
N,
noyau - S, sécrétion ; Sh, spermatophore. Flèches: muscles de la
spermathèque (© A.Lopez C.H.) |
||
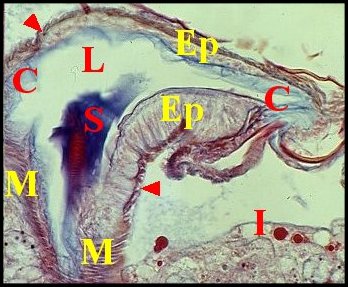 |
| Fig.5- Telema : spermathèque
et
ses muscles pariétaux |
| C,
cuticule - Ep, épithélium - I,
intestin - L, lumière - M, muscles pariétaux en coupe
tangentielle - S, sécrétion. Flèches : sections
transversales
des mêmes muscles (© A.Lopez C.H.) |
En
revanche, la partie postérieure
de la spermathèque a une structure
très différente. Elle s’épaissit
en un disque fortement convexe, épais
de 70 µm et présentant l’aspect d’un
“coussinet”
à priori glandulaire.
A son niveau, la cuticule
s’amincit
considérablement (1 µm) ; l’épithélium
sous-jacent est formé de cellules
très
hautes,
plus sombres, basophiles,
striées
dans leur partie basale et renfermant des noyaux
allongés.
Chez Apneumonella sp.,
la spermathèque
est également
impaire, médiane, sous-ovarienne et présente un aspect
en « besace » très
volumineuse, étendue vers l’avant mais surtout en direction caudale où
son extrémité postérieure décrit une courbe à concavité inférieure. Il
semble bien exister aussi à ce niveau un
épaississement en « coussinet » de l’épithélium pariétal et de la cuticule sus-jacente, jaunâtre et
d’aspect
rigide. Partout ailleurs la paroi, difficile à reconnaître,
est beaucoup plus mince, avec une fine cuticule
sinueuse, un épithélium
aplati
endothéliforme et paraît dépourvue de la tunique
musculaire existant chez Telema. (Fig.6 à 8).
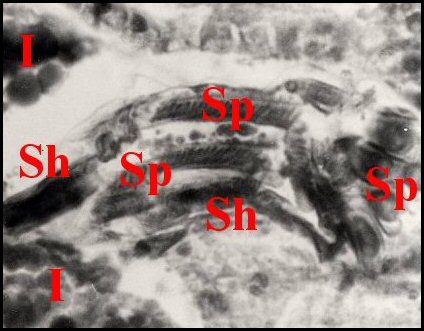 |
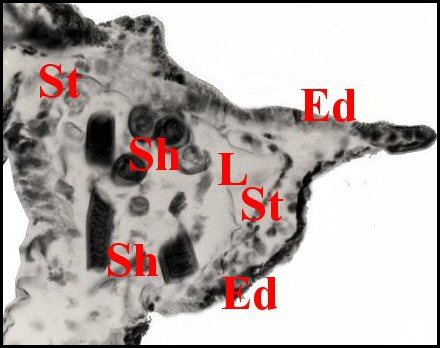 |
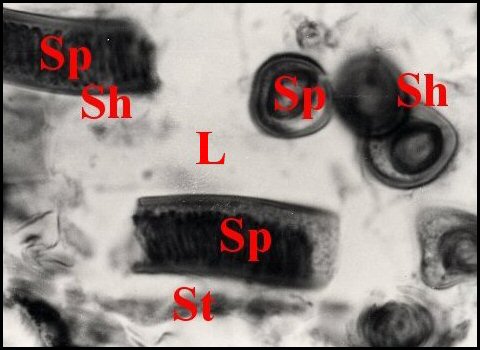 |
| Fig.6-Apneumonella
sp.: spermathèque, coupe en
long |
Fig.7- Autre coupe,
oblique |
Fig.8-
Détail : paroi de la spermathèque et spermatophore |
| Ed, épiderme -
I, diverticule intestinal - L, lumière de la spermathèque - Sh, spermatophore -
Sp, spermatozoïdes
empilés - St, paroi de la spermathèque (© A.Lopez C.H.) |
||
L’unique
femelle observée s’étant accouplée, la cavité renfermait une substance
homogène amorphe et surtout,
un grand spermatophore
jaune-rougeâtre où des gamètes
s’ empilent « comme des comprimés
dans leur étui ». Il diffère toutefois de celui de Telema par son aspect
cylindrique, avec une
cavité axiale régulière et
l’absence complète de digitations (Lopez,1978).
Elle est formée par une basale
uniforme, finement grenue, assez lâche et sinueuse, par
une musculeuse
surtout corporéale,
par une cuticule
et par un épithélium ces deux derniers de
caractères différents au niveau du coussinet
et du corps.
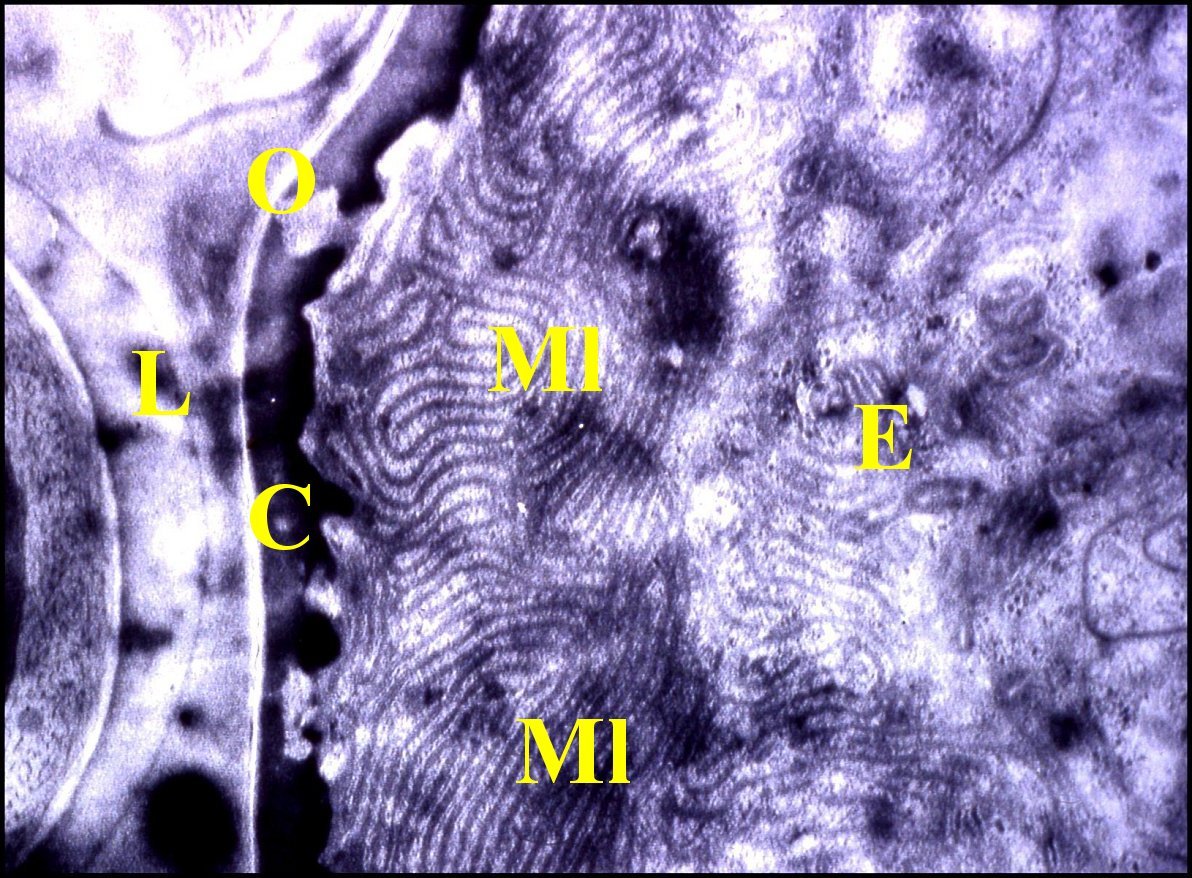
Cet
épithélium est
formé par une seule couche
de cellules.
Leur pôle apical montre des
invagination du plasmalemme
en
replis parallèles, profonds et étroits découpant dans le cytoplasme des expansions très aplaties,
lamellaires, ne
montrant
jamais les contours arrondis de microvillosités
dans les sections
transversales. Ces
expansions ou
microlamelles,
orientées
selon le grand
axe des compartiments cellulaires d’épaisseur
uniforme (50 nm environ), sont jointives, forment une bande continue et
s’y groupent en faisceaux sinueux, plus ou moins incurvés selon leur
longueur (Fig.9). Le cytoplasme
ainsi découpé ne renferme pas
d’autres organites jonctionses.
Il existe des jonctions
solidarisantes à type de desmosomes.
 |
|
|
| C,
cuticule - E, cellules épithéliales - L, lumière - Ml,
microlamelles - O, orifice cuticulaire ( voir flècheFig.10) (© A.Lopez M.E.T.)
|
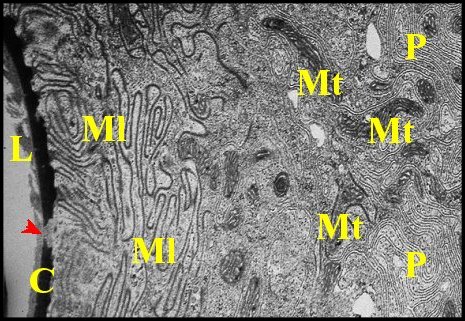
Les deux tiers basaux présentent, de leur côté, un extrême découpage du cytoplasme qui est compartimenté en lanières subparallèles étroites par des replis très profonds du plasmalemme et a donc un aspect lacinié. Ces compartiments semblent accolés les uns aux autres par leurs faces latérales et solidarisés par les jonctions sub-mentionnées. Leur extrémité libre, plus ou moins massuée, renferme parfois une densification du hyaloplasme. Les pôles basaux ainsi conformés rappellent ceux de podocytes.
Le noyau est situé dans la moitié basale de la cellule dont il occupe l’un des compartiments, très élargi à son niveau. Allongé et de contours irréguliers, il renferme de la chromatine en petites mottes et un nucléole volumineux.
Le chondriome est représenté, dans les deux tiers basaux des cellules par de grosses mitochondries plus ou moins ovoïdes, pourvues de crêtes parallèles très serrées et orientées selon le grand axe des compartiments. En revanche, dans le tiers apical, elles sont petites, flexueuses et orientées diversement (Fig.9).
Des organites considérés comme grains de sécrétion sont peu nombreux et situés parfois au contact des mitochondries ; ils ont un contenu homogène et de faible densité.
Le hyaloplasme renferme également des ribosomes libres ou groupés en polysomes.
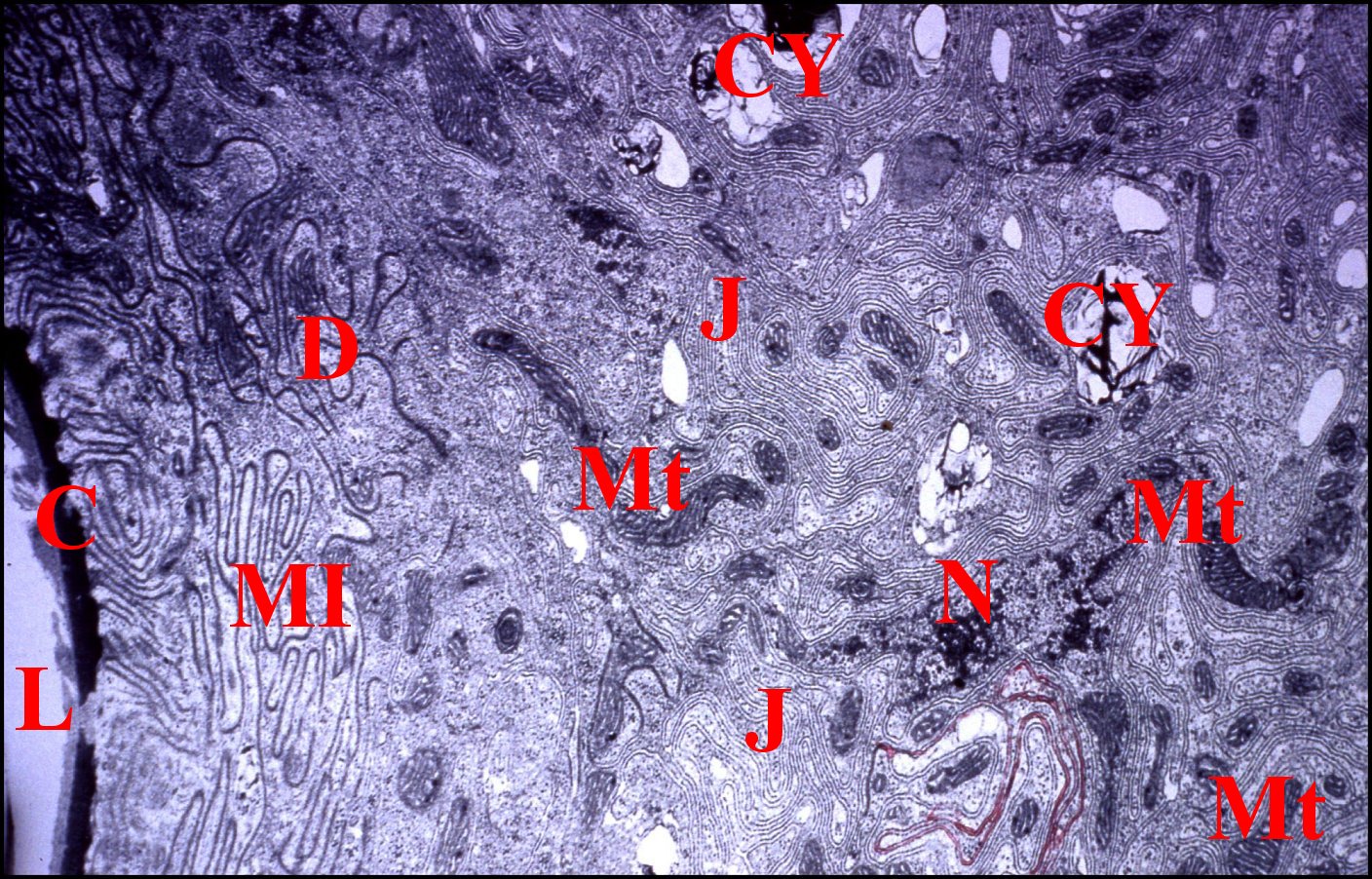
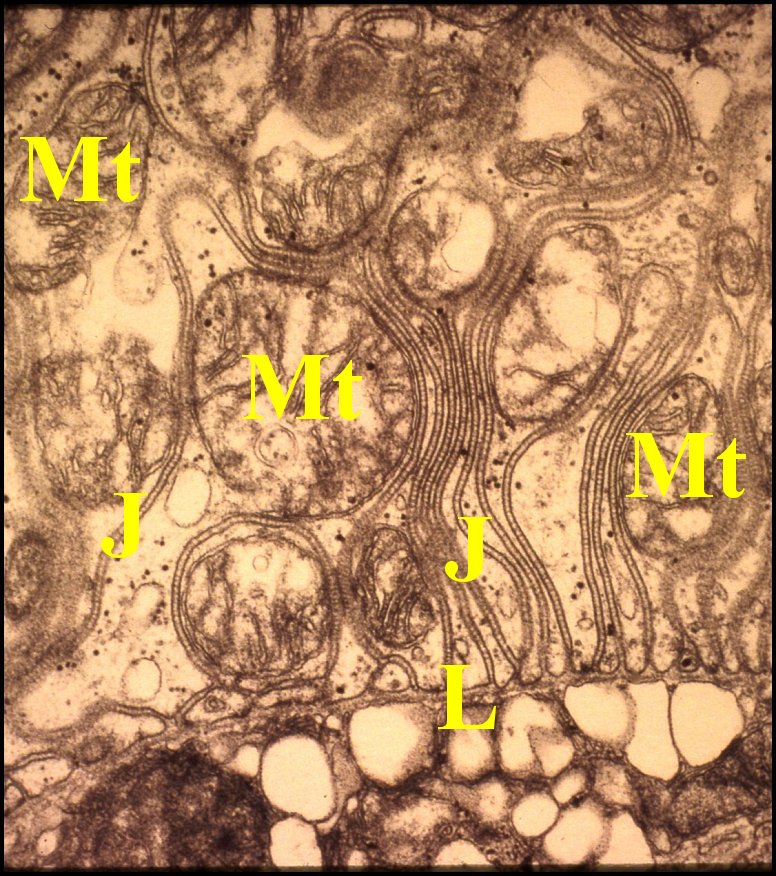
La cohésion des cellules épithéliales est assurée, sur les faces latérales, par des jonctions septées extrèmement nombreuses et spectaculaires (Fig.12 à 14) que des desmosomes zonaires prolongent vers l’apex. Elles se retrouvent au niveau des compartiments des pôles basaux laciniés dont elles solidarisent les franges et rappellent les jonctions septées que possède, au même niveau, la glande coxale sensu-stricto. Du muscle strié est plaqué contre la lame basale ténue
.
 |
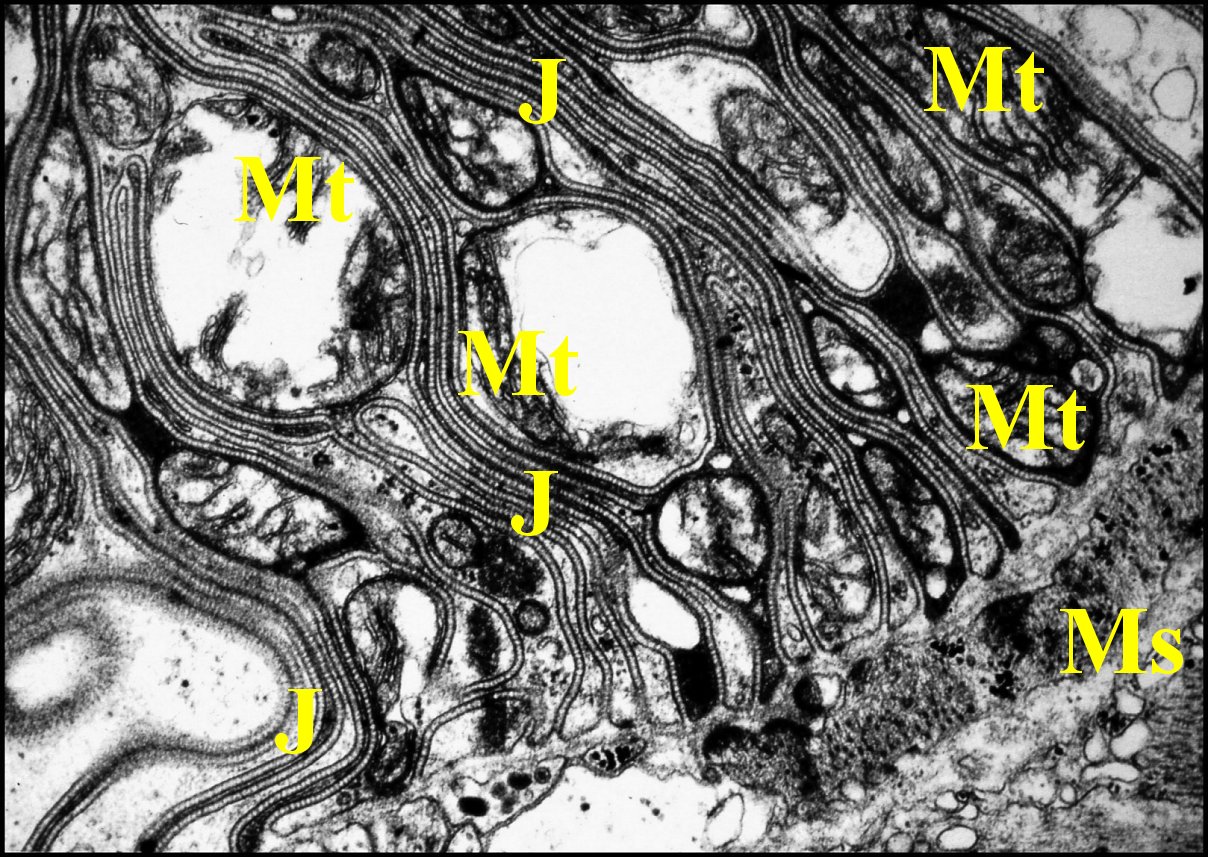 |
|
Fig.12
- Telema
tenella : pôles basaux des cellules épithéliales du
coussinet de la spermathèque.
|
Fig.13-
Telema
tenella : autres pôles basaux des cellules épithéliales du
coussinet de la spermathèque et muscles sous-jacents.
|
|
J,
jonctions septées - L, lame basale - Ms, muscles striés - Mt,
mitochondries (©
A.Lopez M.E.T.)
|
|
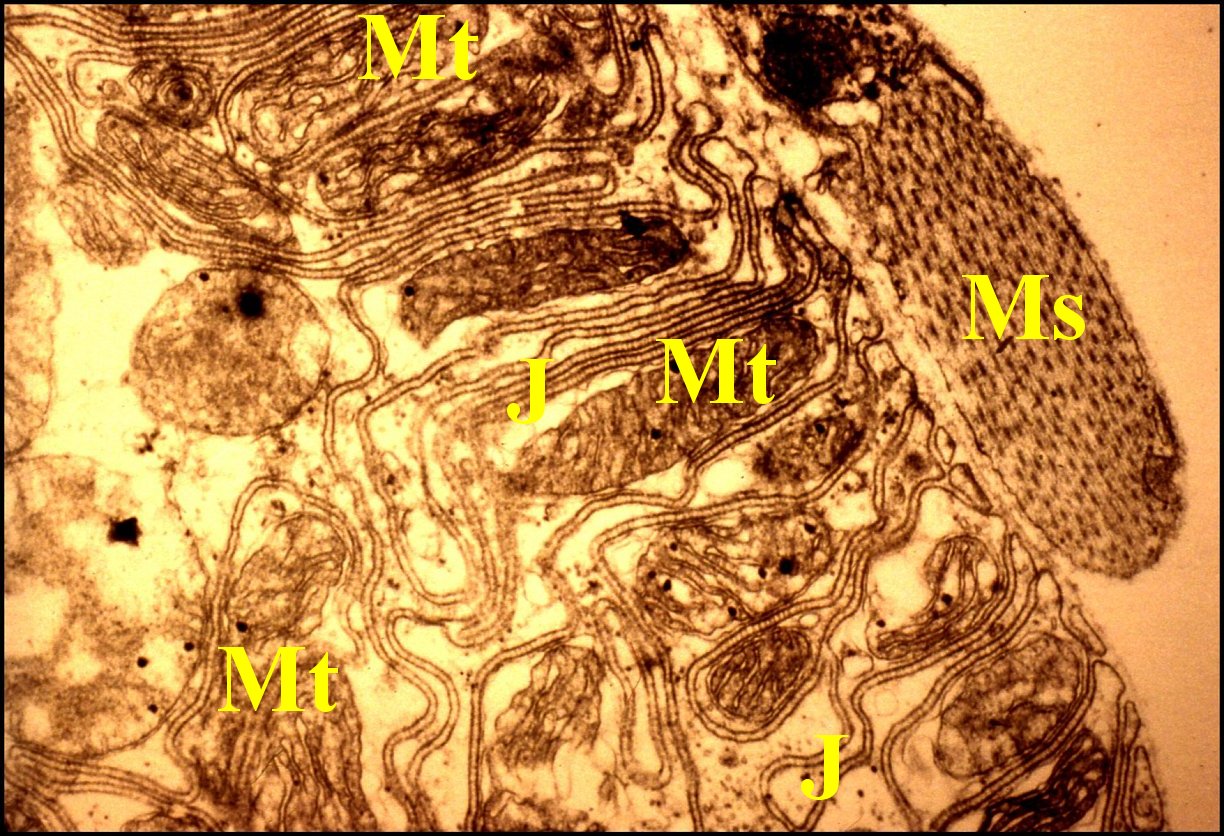 |
|
J,
jonctions septées - L, lame basale - Ms, muscle strié plaqué contre la
lame basale - Mt, mitochondries (© A.Lopez M.E.T.) |
3.1.1.b
-Cuticule du
“ coussinet ”
Elle est
uniquement
épicuticulaire, forme une
couche dense aux électrons et a une épaisseur
moyenne de 250 nm (Fig. 9,10,11). Sa face interne est lisse tout au
long de la lumière tandis que sa face
externe présente une succession de crêtes
plus ou moins arrondies, parfois bifides et se moulant étroitement sur
les expansions lamellaires
sous-jacentes.
Elle est constituée d'une épicuticule externe,
d'une épicuticule interne et
est
traversée par
des pores en forme
d'entonnoir (Fig.10,11).
Ces pores sont en effet plus
larges du
côté
externe que de celui de la lumière ; ils
y
permettent vraisemblablement le passage de substances sécrétées par les
cellules épithéliales.
3.1.2- Corps de la spermathèque
Il est formé par des cellules
jointives plus ou moins hautes. Leur pôle
basal n’est
que légèrement découpé. L’apex
est garni
de villosités
courtes et diversement orientées. Le
noyau
est plus ou moins aplati et pourvu de mottes
chromatiniennes périphériques. Le cytoplasme,
assez dense aux électrons, contient de petites mitochondries
et une grande quantité de corps
pseudo-myéliniques (Fig. ). Le passage de cet épithélium
à celui du coussinet est
assez brusque.
Quelques cellules seulement
présentent
des caractères mixtes.
3.1.2.b- Cuticule
3.1.2.c- Espace sous-cuticulaire
Les cellules épithéliales sont surmontées par un large espace les séparant de la cuticule sus-jacente (Fig.13). Il renferme des corps pseudo-myéliniques libérés, un matériel finement grenu et des masses irrégulières plus denses, dépourvues de membrane .
La
musculeuse
est formée
par une ou deux couches de fibres
striées.
Ces
cellules adoptent une
disposition
annulaire au
niveau du corps de la spermathèque, plus
variable
au niveau du coussinet où elles
se
raréfient (Fig.14). Entourées par des
expansions de la lame basale,
elles
renferment un
noyau marginal assez
irrégulier, quelques
mitochondries et
des myofibrilles
longitudinales.
Ces dernières
convergent parfois vers
des densifications osmiophiles
du myoplasme aux points de
contact avec la basale (Fig.14).
3.2-Contenu
de la spermathèque
Chez
les femelles fécondées, la
lumière du corps
de la spermathèque
contient un spermatophore
baignant dans du matériel amorphe.
Ce
spermatophore est
replié sur lui-même,
agencement remarquable pouvant être déduit des coupes histologiques.
A
partir d’une extrémité, il se
reploie de 3 à 5 fois en « épingles à
cheveux » successives, les replis ébauchant une symétrie rayonnée avec 6
sections en coupe transversale.
De
plus, ils s’opposent alternativement, face ventrale (ou digitée) contre
face ventrale, et face latérale
contre face latérale, les deux
extrémités se trouvant
ainsi du même côté. Une
telle disposition permet au spermatophore d'occuper un minimum de place
dans la spermathèque malgré son volume
relativement
considérable. Il est vraisemblable
qu’elle existe aussi déjà dans le bulbe
copulateur du mâle avant
l’éjaculation.
Comme
dans les coupes
histologiques où son aspect
général demeurait inchangé, le spermatophore conserve son
ultrastructure fondamentale caractéristique . 3 couches superposées
forment toujours sa paroi et ne
présentent que des modifications
minimes (Fig.13) : la couche interne paracristalline s’est un peu épaissie ; les
alvéoles de la couche moyenne se sont élargies ; les
étroits piliers
parallèles de la couche
externe semblent
s’être
inclinés à 45° sur le grand axe
du spermatophore.
Quant aux spermatozoïdes, ils sont moins condensés, de
contour plus régulier, et présentent une coque moins nette, vermiculée
dans la
digitation.
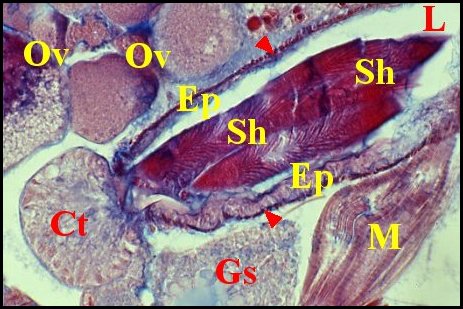
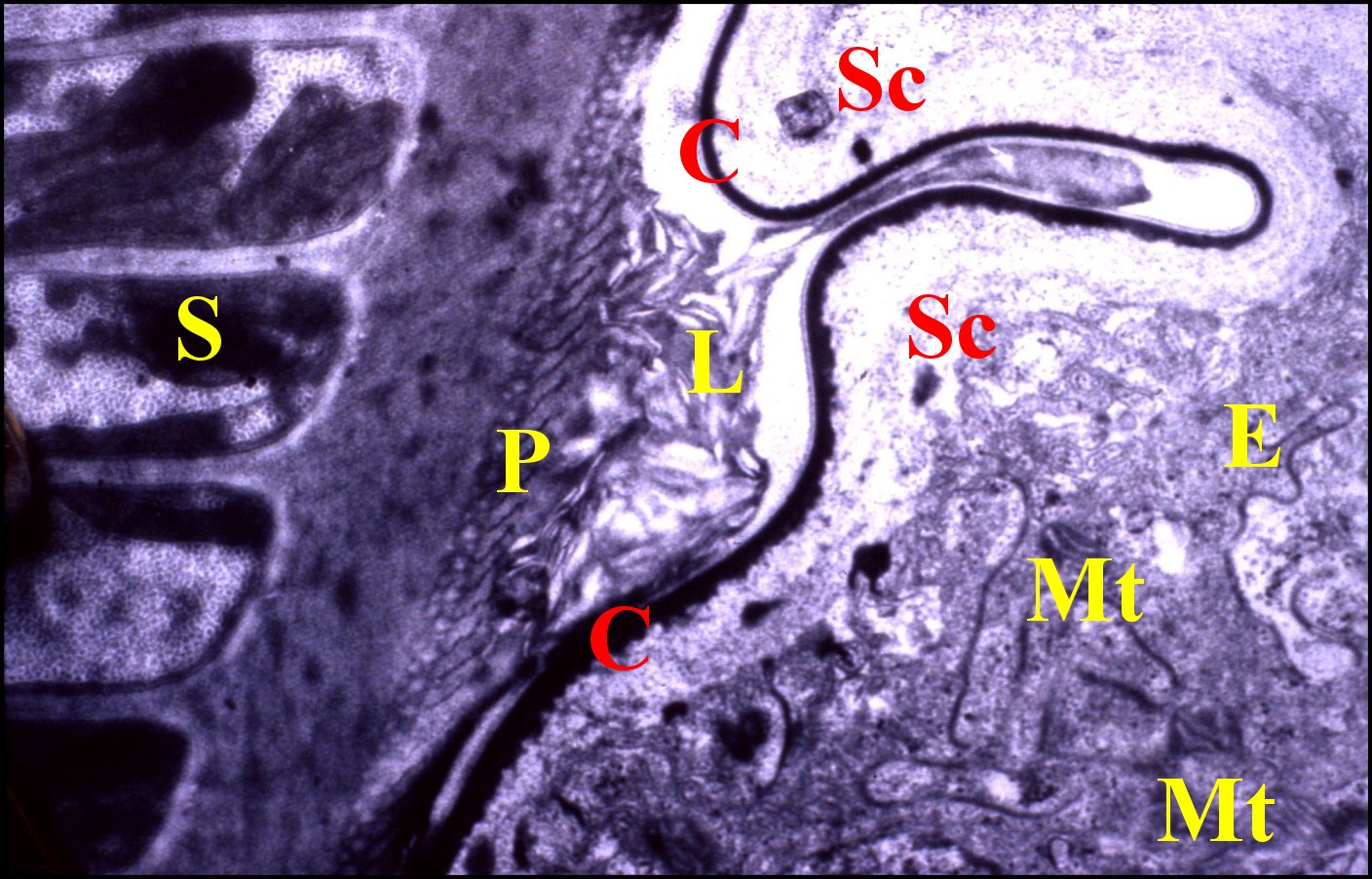 |
| Fig. 13 -
Corps de la spermathèque
contenant un spermatophore dans sa lumière |
| C, cuticule - L, lumière - Mt, mitochondries - Sc, espace sous-cuticulaire - S, spermatozoïdes empilés (© A.Lopez M.E.T.) |
4 -Commentaires
Au
point de vue anatomique, la spermathèque de Télémide
est remarquable
par son volume, si l’on tient
compte de la très petite taille (1,5 mm) d’une femelle de Telema tenella.
Ses dimensions sont en rapport avec celles du spermatophore, introduit dans sa cavité
lors de la copulation et pouvant se replier jusqu’à 5 fois sur
lui-même en «épingles à cheveux» successives pour s’y loger tout
entier (Lopez,1980a).
La spermathèque se
présente comme une simple invagination de l’épiderme au niveau de l’atrium
génital. Sa structure est très
simple si on la compare à celle
d’autres Araignées
«Labidognathes» considérées
comme plus
évoluées, Dysderidae
comprises…
L’étude
ultrastructurale confirme que le coussinet postérieur
est bien de
nature glandulaire et
qu’il s’agit d’une glande anatomiquement définie.
Son épithélium est formé de
cellules sécrétrices toutes
semblables, chacune
d’elles pouvant être assimilée à
une glandule tégumentaire du type 1
tel que
Noirot et (1974,1991) l’ont défini chez les
Insectes. Au même type sont
d’ailleurs rattachées les cellules de la glande rostrale, des
glandes
péribuccales
en «coussinet» ainsi
que celles des conduits de
copulation (Araneus diadematus :
Kovoor,1981).
Toutefois, dans le cas présent, les pôles basaux des adénocytes sont
extrêmement laciniés (découpés en
lanières) et
les villosités apicales classiques
sont remplacées par de curieuses
expansions aplaties formant une bordure en microlamelles trés curieuse.
Le découpage basal des
cellules évoque
celui qui a été noté par
Blest et Taylor (1977
chez d’autres Araignées
du genre Mynoglenes dans
leur prétendue « glande
clypéale ». Elle n’a en
fait aucun
rapport avec le véritable organe du même nom rebaptisée glande acronale par l' auteur,
se
rencontre dans les deux sexes et aurait
plutôt du être appelée
« glande latéro-sousoculaire » . De
plus, ses cellules possèdent
des canalicules excréteurs individuels et ont des compartiments
basaux plus irréguliers, renfermant
peu de mitochondries.
La laciniure
basale et la
richesse du chondriome associées dans le coussinet de la spermathèque
sont donc beaucoup plus proches
du labyrinthe de la glande coxale, ce dernier possédant
toutefois des
jonctions
dites scalariformes .
La bordure
apicale microlamellaire est un caractère morphologique
très inhabituel découvert dans les tubes de Malpighi et l'intestin d'Insectes Homoptères (Smith & Littau,1960),
Diptères (Waterhouse & Wright,1960)
ainsi
que chez les Arachnides, dans les tubes excréteurs de l’Acarien Macrocheles muscaedomesticae (Coons
& Axtell,1971). Une
ultrastructure un peu voisine semble d’ailleurs se
retrouver dans
l’ordre des Araignées, où des microvilli apicales « foliacées »
caractérisent la glande
labiosternale des Theridiosomatidae (confere supra).
Au point de vue fonctionnel,
il doit se produire des
échanges extrêmement actifs au niveau du pôle basal comme les mitochondries en témoignent par leur nombre, leur
taille et
leur association avec ses replis membranaires.
La sécrétion des cellules du coussinet
gagne vraisemblablement la lumière
de la spermathèque au travers des pertuis en entonnoir ajourant la cuticule. Son rôle ne peut guère être
précisé mais pourrait bien être lié à l’existence du spermatophore : nutrition des gamètes mâles, lyse de leur coque d’enkystement et de la paroi spermatophorique, entrainement
mécanique vers les ovules à féconder
pendant la ponte.
Les déplacements du spermatophore
lui même pourraient être facilités par les contractions de la musculeuse, ainsi que par les
modifications structurales de la couche
externe dont les piliers (Lopez,1980a ; Lopez,1980b)
subissent une inclinaison (Fig. ) après qu’il ait été
introduit dans la spermathèque (Lopez,1983a : fig.5a,5b ; Lopez,1983b).
Au
point de vue
systématique, il est probable qu’une même
structure de la spermathèque est commune à toutes les Araignées
Telemidae si
l’on en juge par la description qu’en ont fait divers auteurs (Saaristo,1978 ;
Brignoli,1978,1980 ; Baert,1980) d’après les préparations
éclaircies de genres différents (Seychellia, Cangoderces, Usofila, Jocquella).
Au
point de vue
phylogénique,
la structure
de la spermathèque
distingue nettement Telema
tenella,
Apneumonella
et probablement aussi toutes
les autres Telemidae,
des LeptonetidaeFig. ). De
même, elle ne paraît pas autoriser un rapprochement avec les Dysderidae bien qu’il existe quelques
ressemblances dans les glandes coxales (Lopez,1983d)
et séricigènes (Lopez,1983c).
Note 1 : Laboratoire CNRS, Moulis 09200 : fixation des parties étudiées au glutaraldéhyde à 2,9% dans le tampon Millonig 0,2 M, post-fixation au tétroxyde d’osmium à 0,2% dans le même tampon, et inclusion en épon ; coupes fines au microtome Reichert OM U2 contrastées par l’acétate d’uranyle, le citrate de plomb et examinées ensuite sous 50 KV, au microscope Sopelem du Laboratoire souterrain. Travail effectué avec Lysiane Juberthie-Jupeau, directeur de recherches au CNRS, et ses collaboratrices.
Bibliographie
Baert,L.,1980.-
Bull.Br.Arachnol.Soc.,5,1,p.16-19.
Blest,A.D.
&
H.H.Taylor,1977.- J.Zool.Lond.,183, p.473-493.
Brignoli,P-M., 1978.-
Rev.Suisse Zool.,
85,1, p. 111-114.
Brignoli,P-M.,
1980.-
Rev.Suisse Zool.,
87,2, p. 605-609.
Coons,L.B.
&
R.C.Axtell,1971.- J.Morph.,133, p.319-338.
Fage,L.,
1913.- Arch.Zool.exp.gen.,10
(Biospeologica ,29), p.479-576.
Kovoor,J.,
1981.-
Atti.Soc.Tosc.Sci.nat.,Mem.ser.B,788, suplt.p.1-15.
Lopez,A,1976
(avec H.Salvayre).- Bull.Soc.Et.Sci.nat.Béziers,
N.S., IV, Vol.45, 1976, p.17-26.
Lopez,A.,1977.-
Bull.soc.zool.France, 102, n° 3, p.261-266.
Lopez,A.,1978
(avec R.Legendre).- Bull.soc.zool.France, 103, n° 1, p.35-41.
Lopez,A.,1980a
(avec C.Juberthie).- C.R.Veme Coll.Arach.,Sept.1979, Barcelona, 1980,
p.111-117.
Lopez,A.,1980b
(avec C.Juberthie & J.Kovoor)– Intern.Journ.Invert.reprod., 3,
p.181-191.
Lopez,A.,1983a.- Bull.Soc.Et.Sci.nat.Béziers,
N.S.,
IX, Vol.50, 1982-1983, p.20-28.
Lopez,A.,1983b
(avec L.Juberthie-Jupeau).- Mém.Biospéol, X, p. 413-418.
Lopez,A.,1983c
(avec J.Kovoor).- Mém.Biospéol, X, p. 419-425.
Lopez,A.,
1983d (avec L.Juberthie-Jupeau &
J.C.Bonaric).- Mém.Biospéol, X, p.433-437.
Noirot,C.
& A.
Quennedey.,1974.- Ann.Rev.Ent., 169, p.61-80.
Noirot,C.
& A. Quennedey., 1991.- Annls
Soc.ent.Fr. (N.S.), 1991, 27 (2), p.123-128.
Saaristo,M.I.,1978.-
Ann.Zool.Fenn.,15, p.99-126.
Simon, E., 1882.- Etudes arachnologiques, 13 eme mémoire. Ann.Soc.ent.France (6) 2, p.204-205
Simon,
E.
& L.Fage,1922.- Arch.Zool.exper., 60, (Biospeologica 46), p.523-555.
Smith, D.S. &
V.C.Littau,1960.-
J.Biophys.Biochem.Cytol.,8, p.103-133.