Glande
clypéale ou
acronale des Argyrodes
|
Les
Araignées Theridiidae mâles du genre Argyrodes
possèdent
une glande exocrine
remarquable située au niveau du clypeus (bandeau), donc dans
la
partie la plus antérieure du corps ou acron et élaborant
une sécrétion qui intervient lors de
l'accouplement. Elle est probablement présente aussi dans les
autres genres d' Argyrodinae
|
| Couleurs
conventionnelles : En noir et italiques, termes anatomiques ; en violet,, noms génériques et spécifiques ; en vert, noms de familles et sous-familles ; en orange,, parties les plus importantes et résumés; en bleu, liens divers. |
|
Abréviations
conventionnelles :
M.E.B. : (photographie en) microscopie électronique à balayage M.E.T. :
(photographie en) microscopie
électronique à transmission
C.H. : coupe histologique (microscopie photonique) |
1 - Introduction
2 - Le dimorphisme sexuel prosomatique acronal2-1 Morphotypes
2-1-1 Les Types
Type acuminé
Type rostré
Type nasuté
Type lippu
Type prognathe
Type camard
2-1-2 Les reliefs tégumentaires
3 - La glande acronale sous-jacente
3-1 - Structure histologique
3-2 - Ultrastructure
3-2-1 - Unité glandulaire
3-2-1a - Adénocyte
3-2-1b - Portion réceptrice (canalicule récepteur)
3-2-1c - Portion conductrice (canalicules conducteurs et leurs cellules)
4 - Commentaires
4-1- Sur le plan anatomique
4-2- Sur le plan fonctionnel
4-3 - Sur le plan systématique
4-3-a - Cas des Argyrodinae
4-3-b- Hors des Theridiidae : Linyphiidae
1 - Introduction
Le clypéus
des
Araignées est un territoire plus ou moins étendu
correspondant à la partie toute antérieure de la face
dorsale du prosoma
(“ carapace ”
ou “ bouclier ”)
(Fig.1).
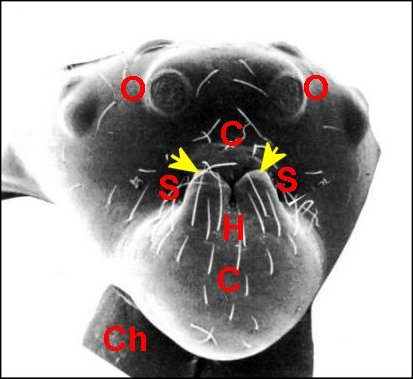
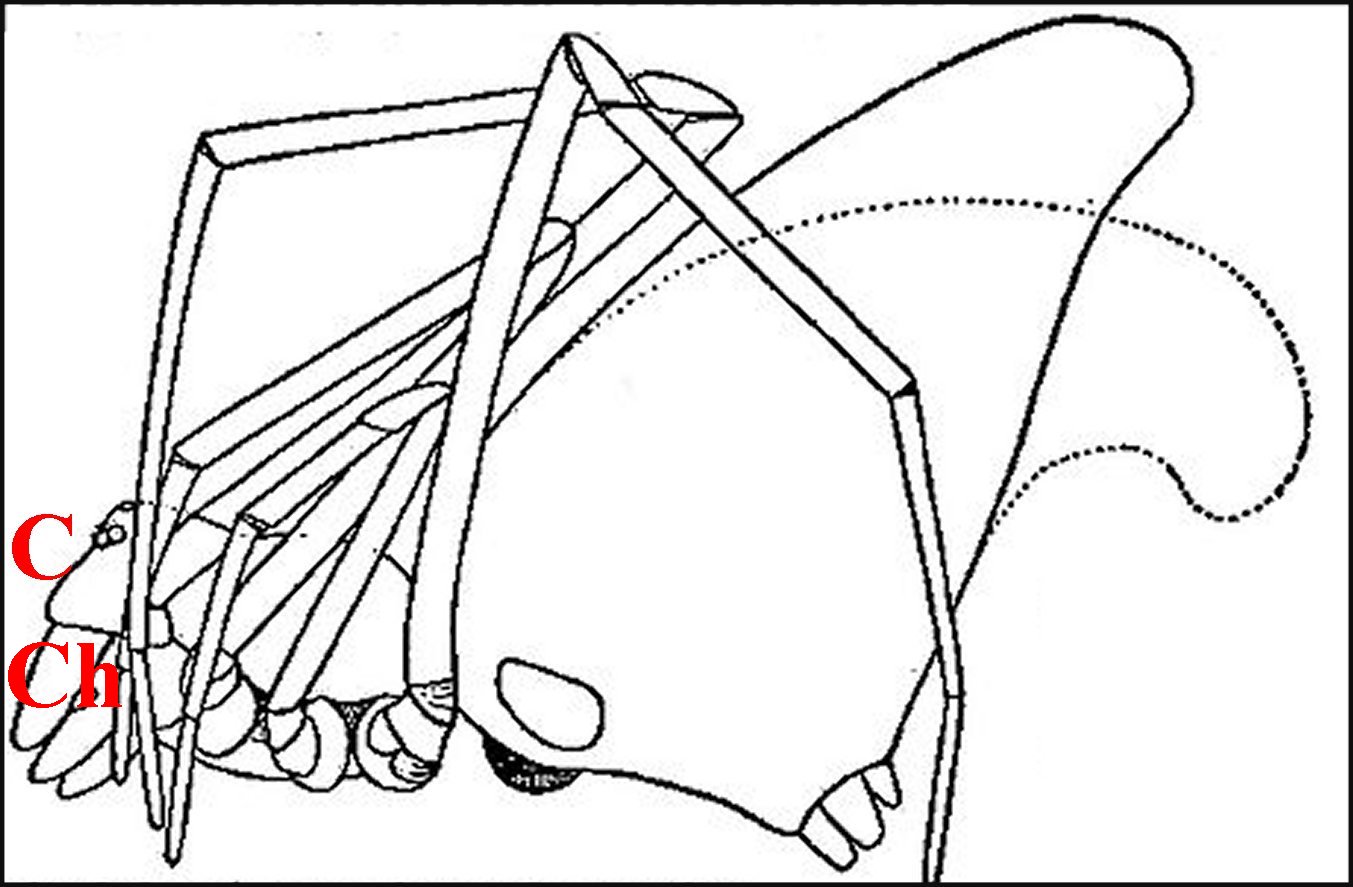
Encore appelé “ bandeau ”, il sépare les yeux du bord frontal (B) surplombant les chélicères. On peut considérer qu’il fait partie de l’acron : région “acronienne” ou céphalique primaire prébuccale.
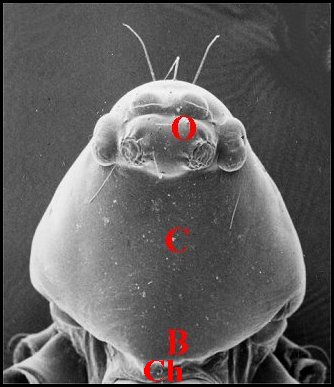 |
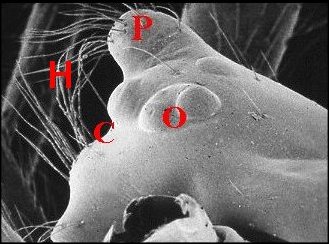
| Fig.1
- Clypeus ou
bandeau (C) particulièrement
développé chez
une Microdipoena
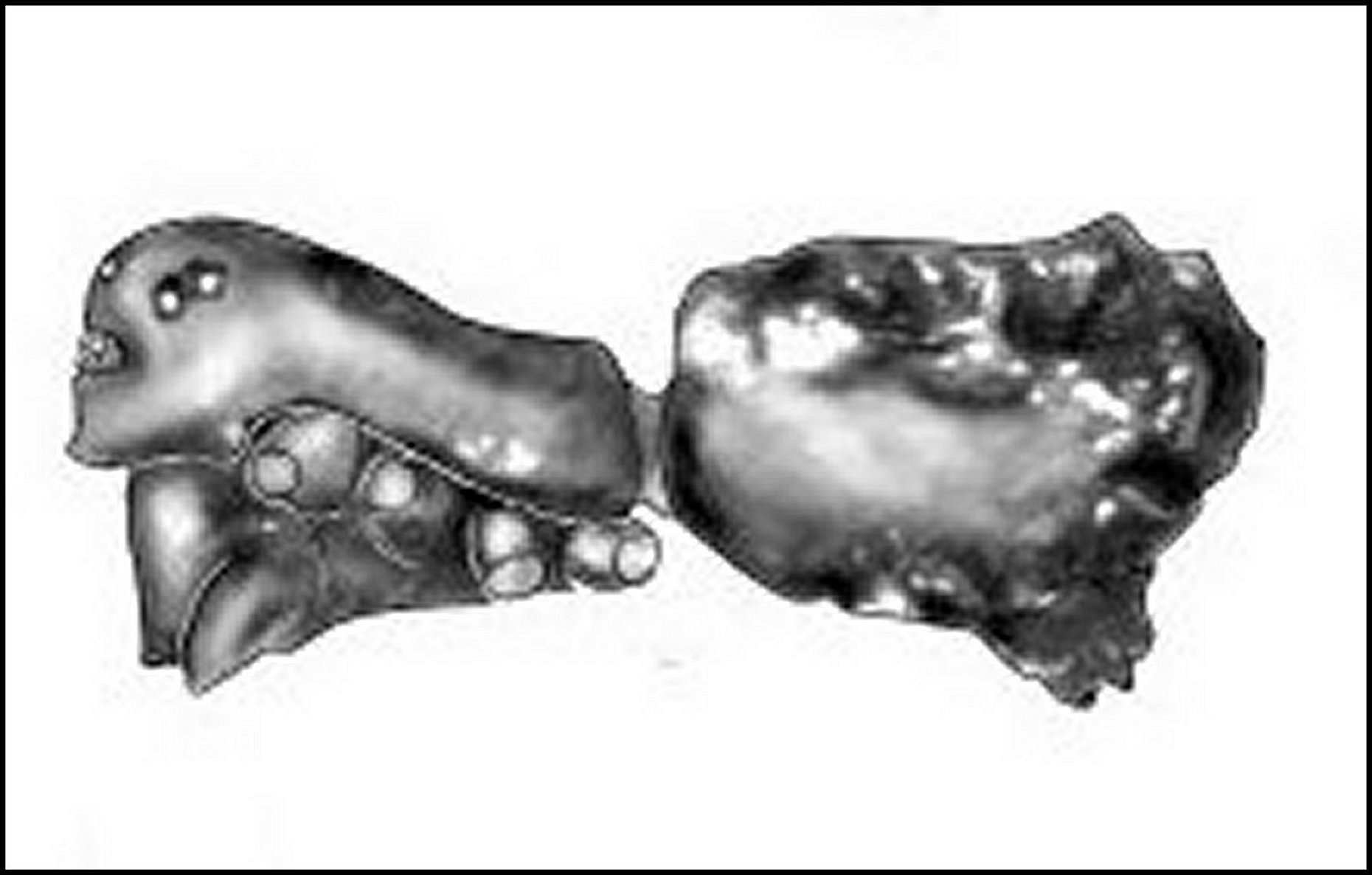
(Mysmenidae) de Mahé, Seychelles |
| B, bord
frontal - C, clypeus
- Ch,
chélicères - O, groupe oculaire (© A.Lopez M.E.B.) |
Chez les mâles de
certaines Araignées
(Theridiidae du genre Argyrodes et Thymoites,
Linyphiidae
Erigoninae), il est en relation profonde avec
un épiderme dont les cellules se sont
différenciées en organes tégumentaires
sécréteurs plus ou moins complexes, les glandes
dites initialement clypéales.
Le même nom leur a été attribué à
tort et par extrapolation par Taylor et Blest (2009) chez
des Araignées Linyphiidae
du genre Mynoglenes
où elles sont en fait bilatérales, bas situées et
surtout sous-oculaires (Fig.2). Leur ultrastructure est en outre fort différente.
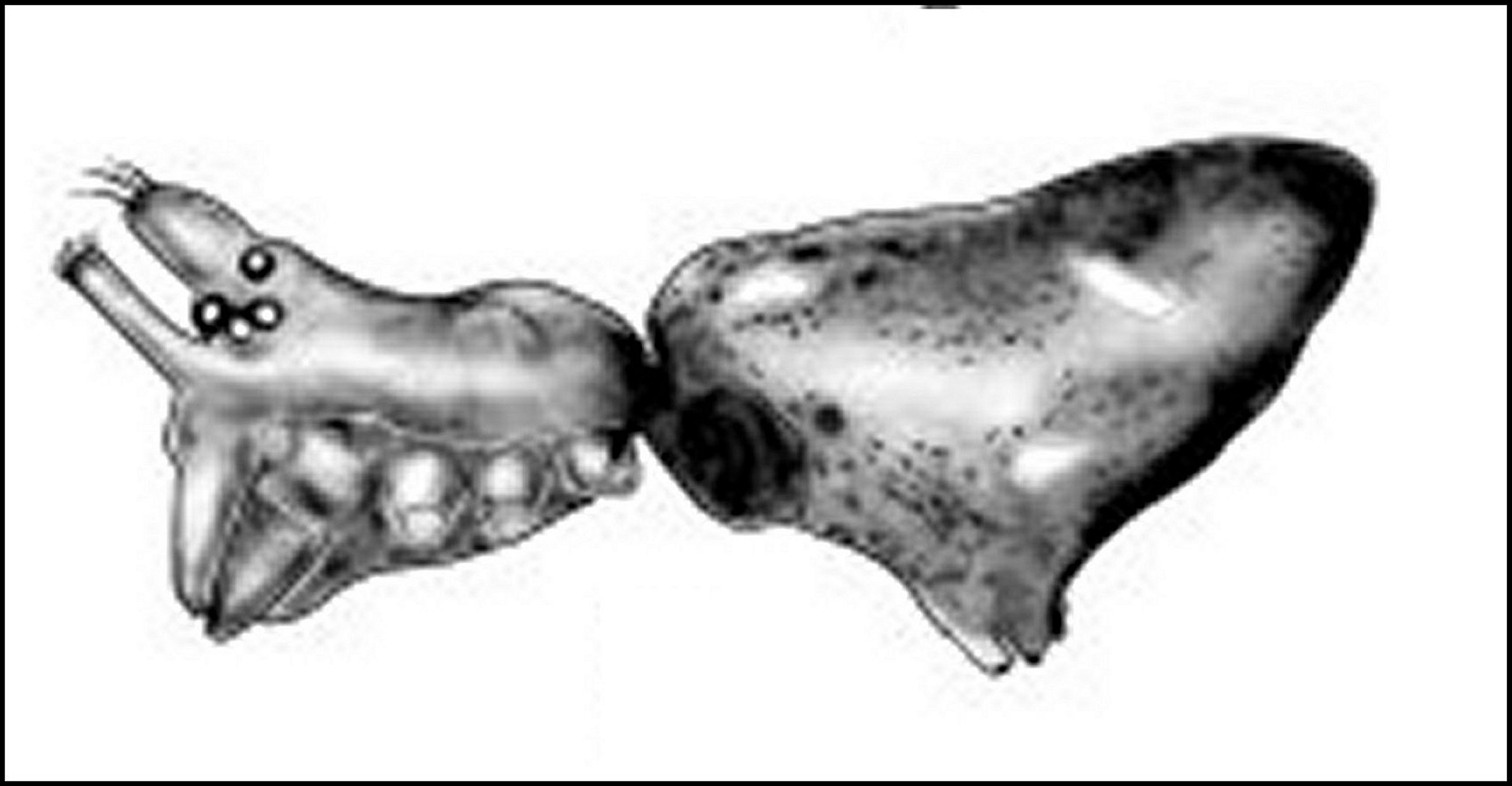
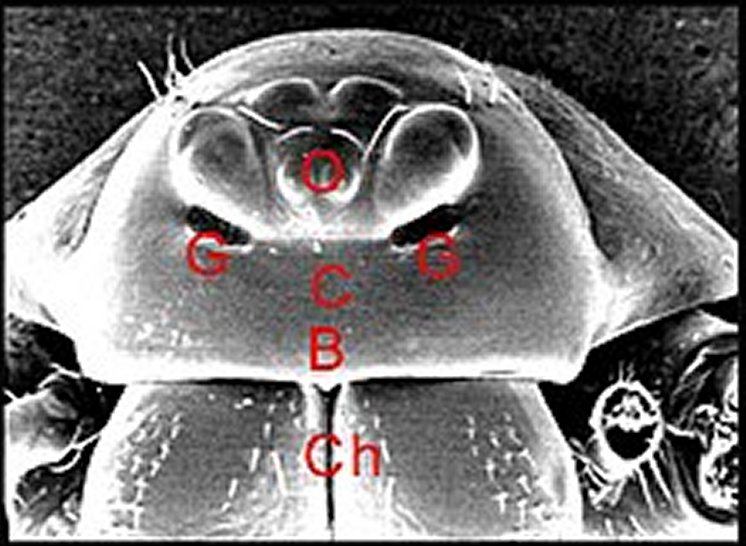 |
| Fig.2 - Vue de face ou frontale
d'une Araignée Mynoglenes
mâle de la Réunion.
|
|
B, bord frontal - C,
clypéus - Ch, tiges chélicériennes -G, orifices des glandes dites
"clypéales", en fait sous-oculaires - O,
région oculaire
(© A.Lopez M.E.B.) |
L'auteur, André Lopez, l'a découverte chez
les Argyrodes il
y a donc aujourd'hui (2023) près de 50 ans,
en examinant les coupes
histologiques de l'une d'entre elles, Argyrodes
zonatus (Walck.), récoltée
à Madagascar (Dr.Rakotondrainibe), retrouvée
ultérieurement dans la même île ainsi qu'à la
Réunion (Lopez,1990) et à
Mayotte (Lopez, 2010) respectivement sur
des toiles de Nephila
inaurata et comorana,
superbes Aranéides tropicales françaises.
Les Argyrodes
sensu stricto sont si
intéressantes par leur comportement et d'autres
caractères anatomiques qu'ils
sont présentés en détail dans un article du site
dédié spécialement au genre et à ses
divers hôtes, en particulier les Nephiles (Nephila
et Trichonephila).
La glande clypéale, organe
prosomatique insolite, est propre aux mâles du genre
Argyrodes,
sans équivalent chez leurs
femelles et remarquable par sa localisation (Lopez,1974a
; Lopez, 1977b).
Il
siège en effet dans la
région "céphalique"
du corps de l' Argyrodes
mâle, plus précisement dans la
partie
antéro-supérieure du prosoma où ses rapports
avec le clypeus
avaient d'abord
inspiré à Lopez et Legendre
(1974) son premier nom de “glande
clypéale”,
consacré jusqu’ici par
l’usage et étendu outrancièrement à d’autres
familles d’Araignées.
En
fait, cette appellation globale
ne paraît plus justifiée à l'auteur car chez
certaines espèces
du type “acuminé” ,
la glande n’a plus, au moins en partie, de relation directe avec le
“ bandeau ”.
Mieux vaut donc rattacher l’organe
à l’acron et le
désigner sous un nouveau nom :
glande
acronale.
Sa
présence est
étroitement liée à celle d’un dimorphisme
sexuel prosomatique
acronal très
marqué.
2 -
Le Dimorphisme
sexuel
prosomatique acronal
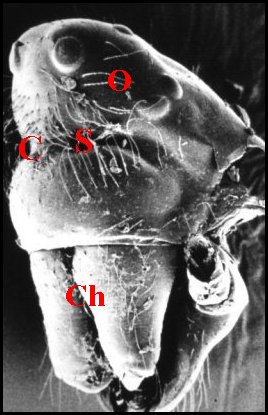
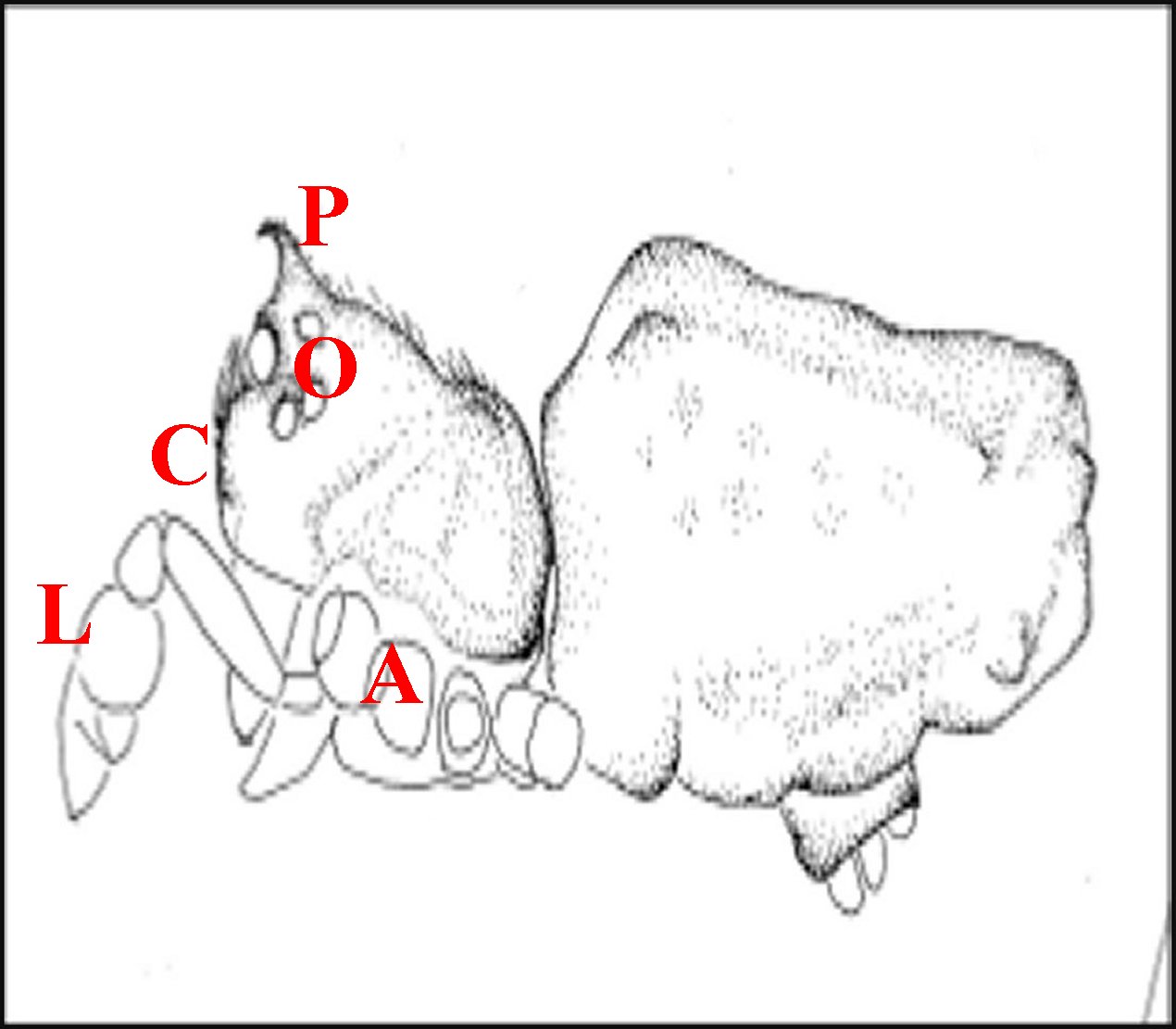
Le prosoma des femelles a un aspect banal et assez uniforme (Fig.3).
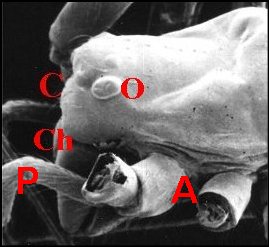 |
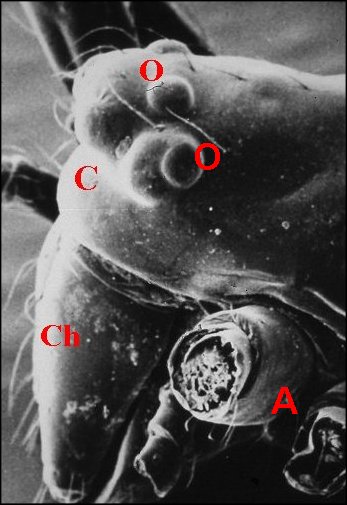 |
|
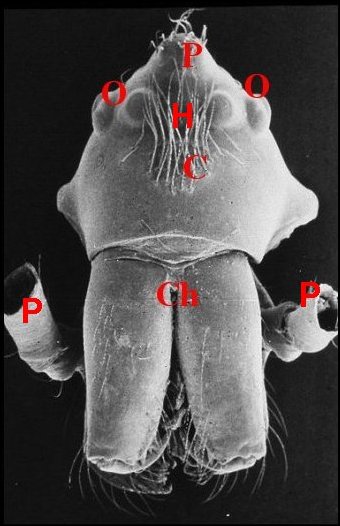
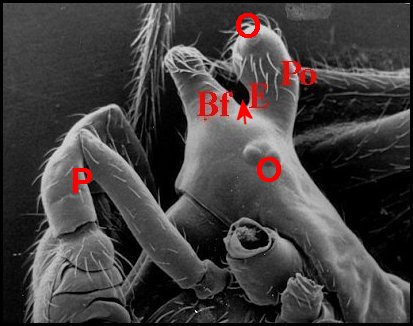
Fig.
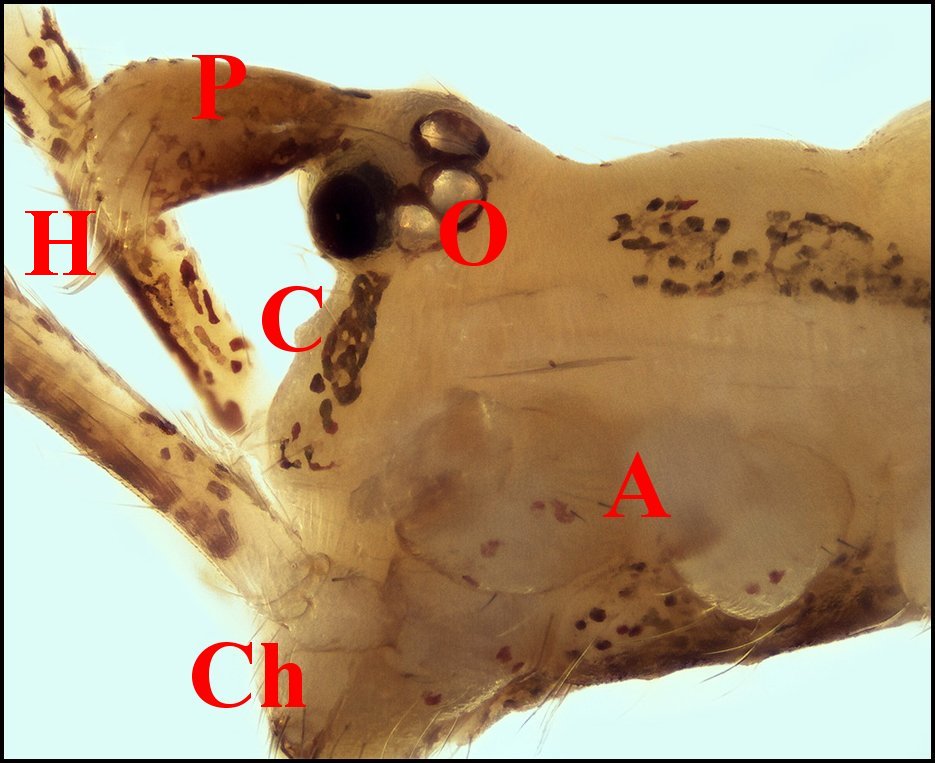
3 - Argyrodes femelles
: A, patte ambulatoire - C, clypeus
d'aspect
banal - Ch, chélicère - O,
oeil - P, pédipalpe (© A.Lopez
M.E.B.)
|
|
En
revanche, celui des mâles présente une morphologie
extraordinaire bien
visible au M.E.B. dans sa région
acronale.
Associant des saillies, bosses ou
protubérances et des dépressions, échancrures ou
sillons, cette
anatomie réalise en quelque sorte une “galerie des monstres”
tant elle est surprenante et peut même “choquer” par sa
présentation comparative chez diverses espèces.
Suivant
l’aspect
général
et la disposition de ces reliefs, l'auteur avait proposé
initialement
de
rattacher les mâles d’ Argyrodes
à quatre types
morphologiques bien distincts,
du moins chez les espèces alors
connues :
types “nasuté”, “rostré”, “lippu”
ou “prognathe”et “camard”
(Lopez, 1977b; Lopez, 1979).
2 -1 En
fait, une nouvelle
classification, comportant cette fois 6 morphotypes,
paraît
plus
appropriée à l'auteur. Tous montrent des reliefs accessoires, des poils et surtout les orifices ou pores excréteurs de la glande acronale.
►Le
nouveau type dit aujourd'hui acuminé se caractérise
par
une
protubérance unique
droite ou procurvée, en "tourelle" ou
en
"cimier", surplombant le groupe
oculaire et inclut ainsi des
espèces
telles qu’ Argyrodes
cognatus
(Seychelles) (Fig.3,4) et A.projiciens (Amérique
tropicale) (Fig.5).
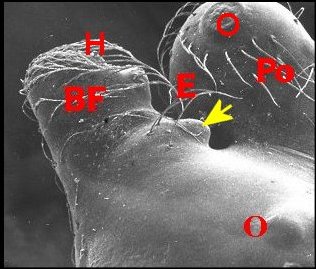
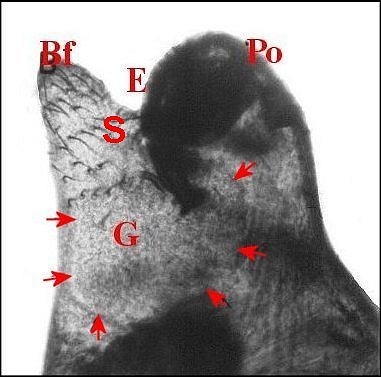
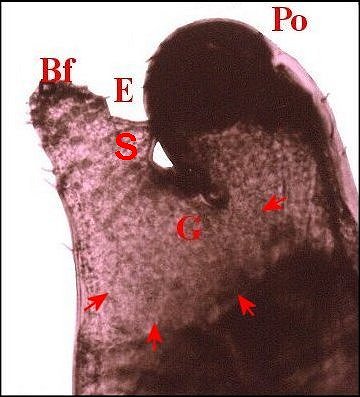
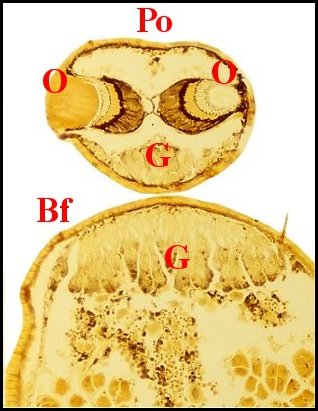
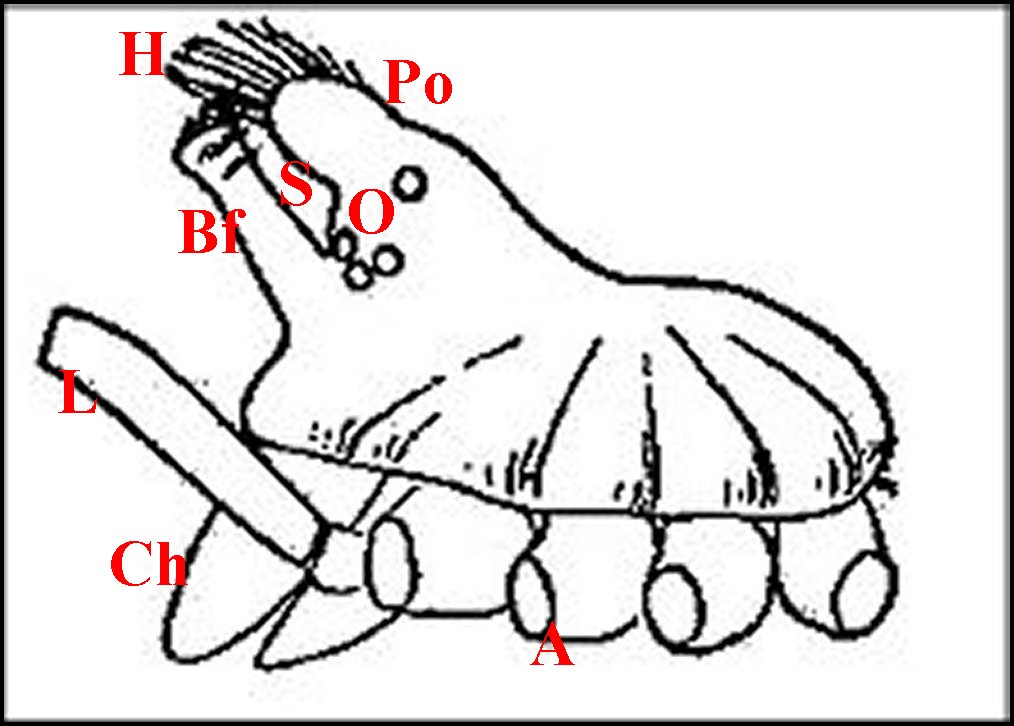
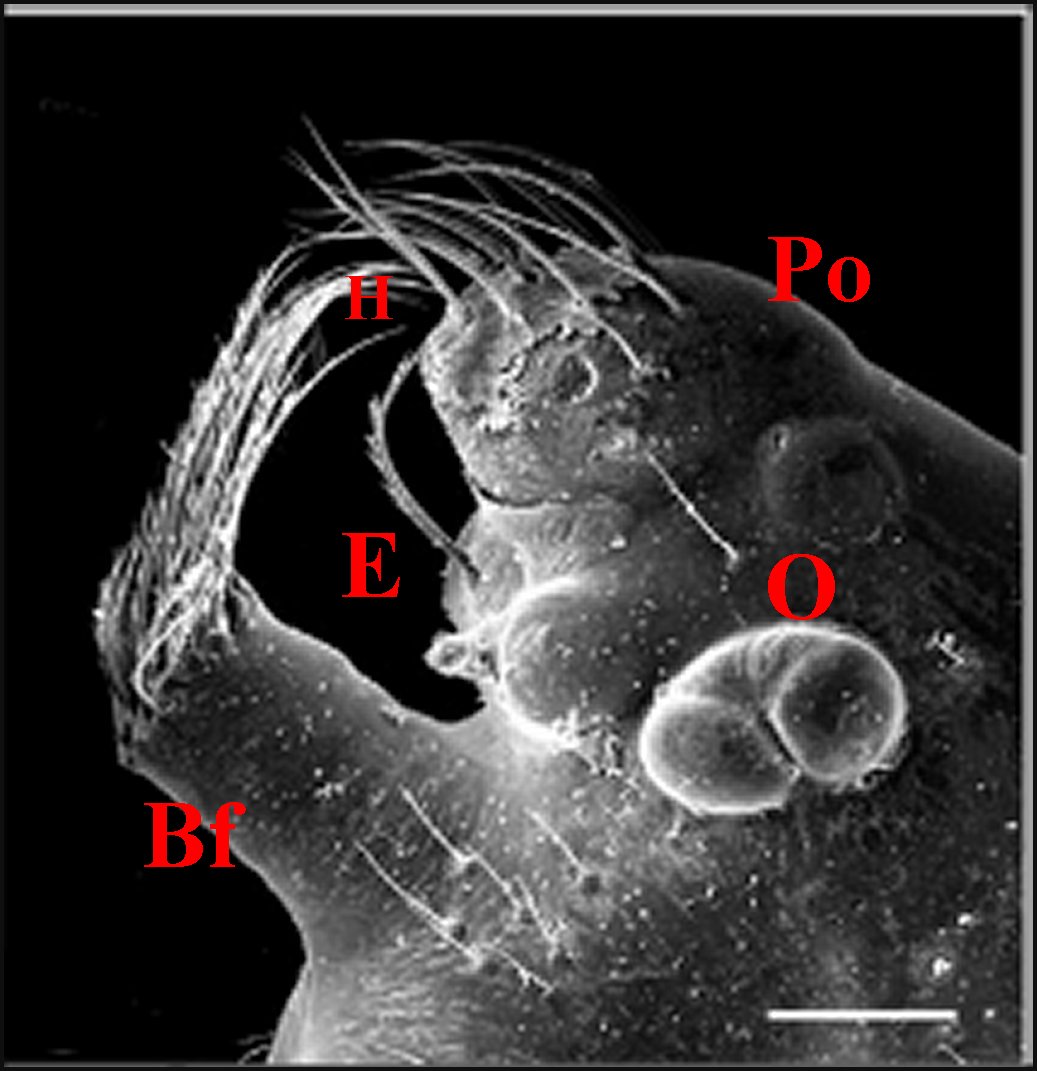
►Le type 2
déja
appelé rostré
(Fig.6,7)
convient
mieux aux mâles
dont le clypeus
est
divisé en deux projections par une échancrure (E) :
la protubérance oculaire
(Po) et la bosse frontale
(Bf),
cette dernière évoquant effectivement un rostre ; il
concerne une
majorité d’espèces, d’abord rattachées au
“ nasuté ”,
notamment Argyrodes
elevatus, de Guadeloupe (Fig.6), A.zonatus, de
Madagascar et autres îles de l' Océan indien (Fig.7), A.nephilae,
de Guadeloupe (Fig.8), A.rostratus
nephilae, des Seychelles (Fig.9)
et le taxon
cosmotropical A.
argyrodes (Fig.22)
|
 |
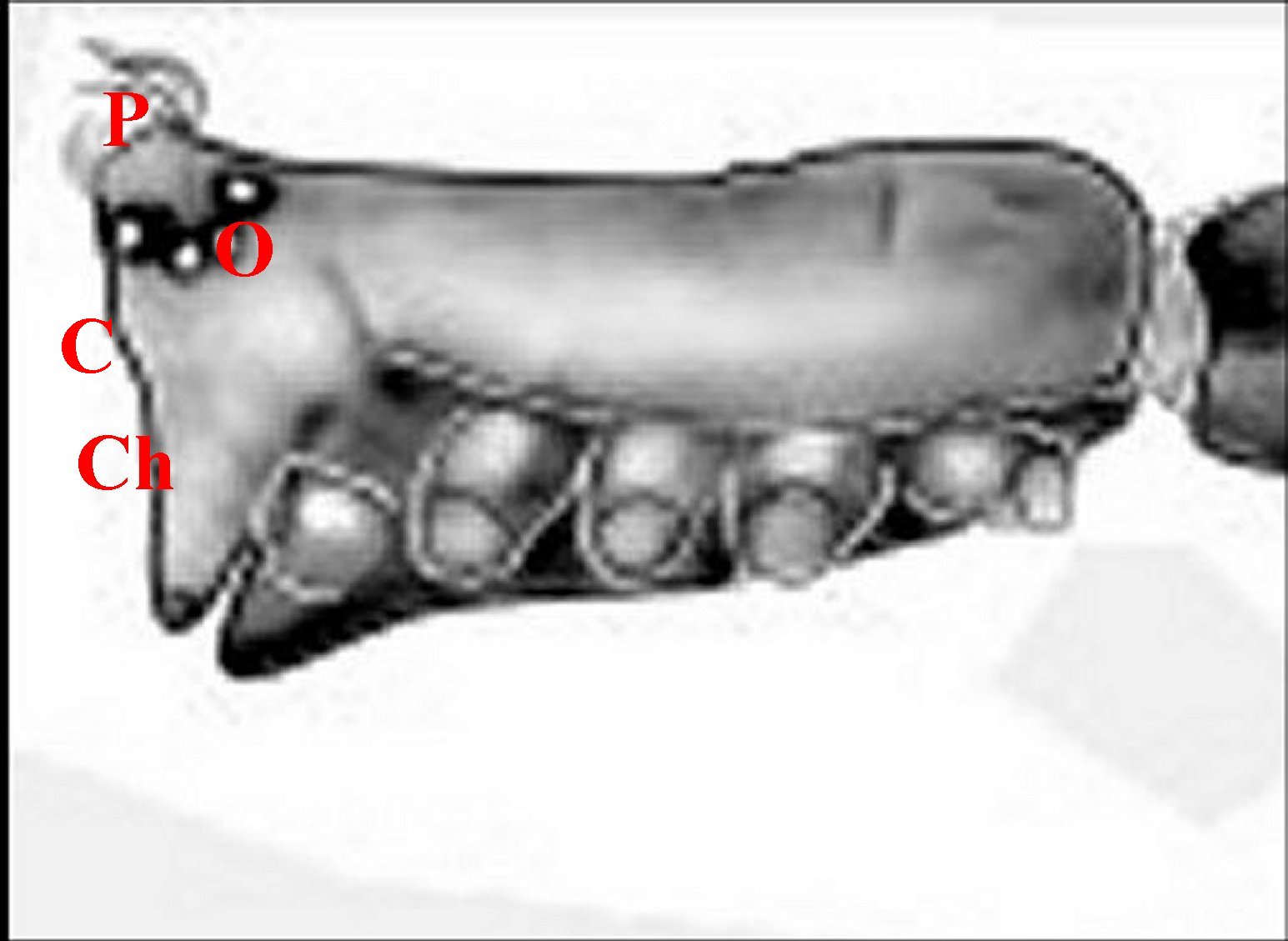
| Fig.6 - Argyrodes
elevatus, vue latérale
gauche |
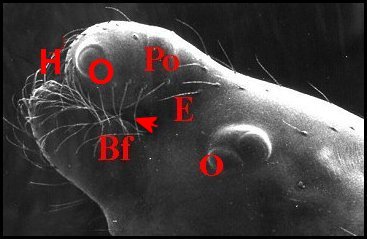
Fig.7 - Argyrodes zonatus, vue latérale gauche |
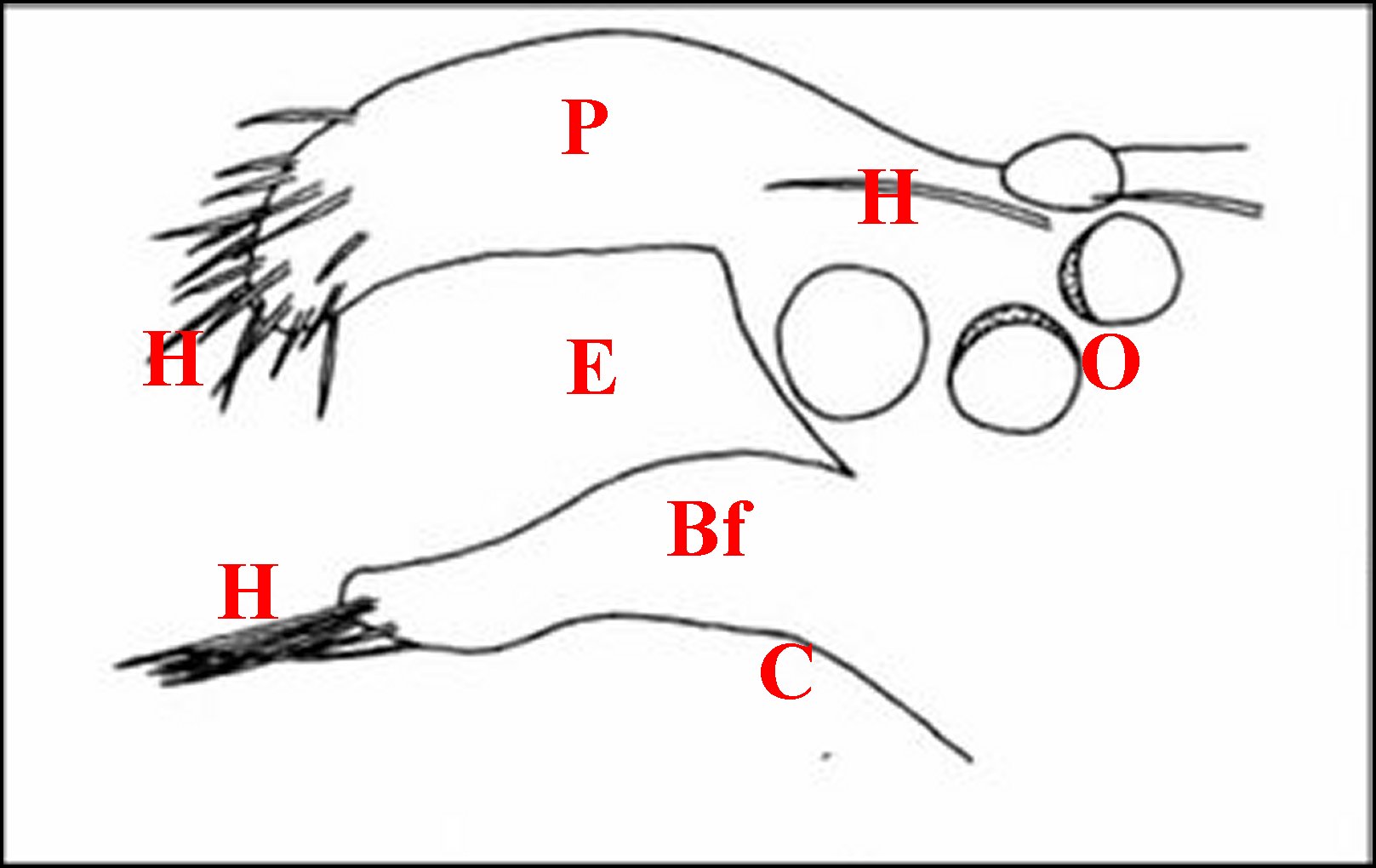
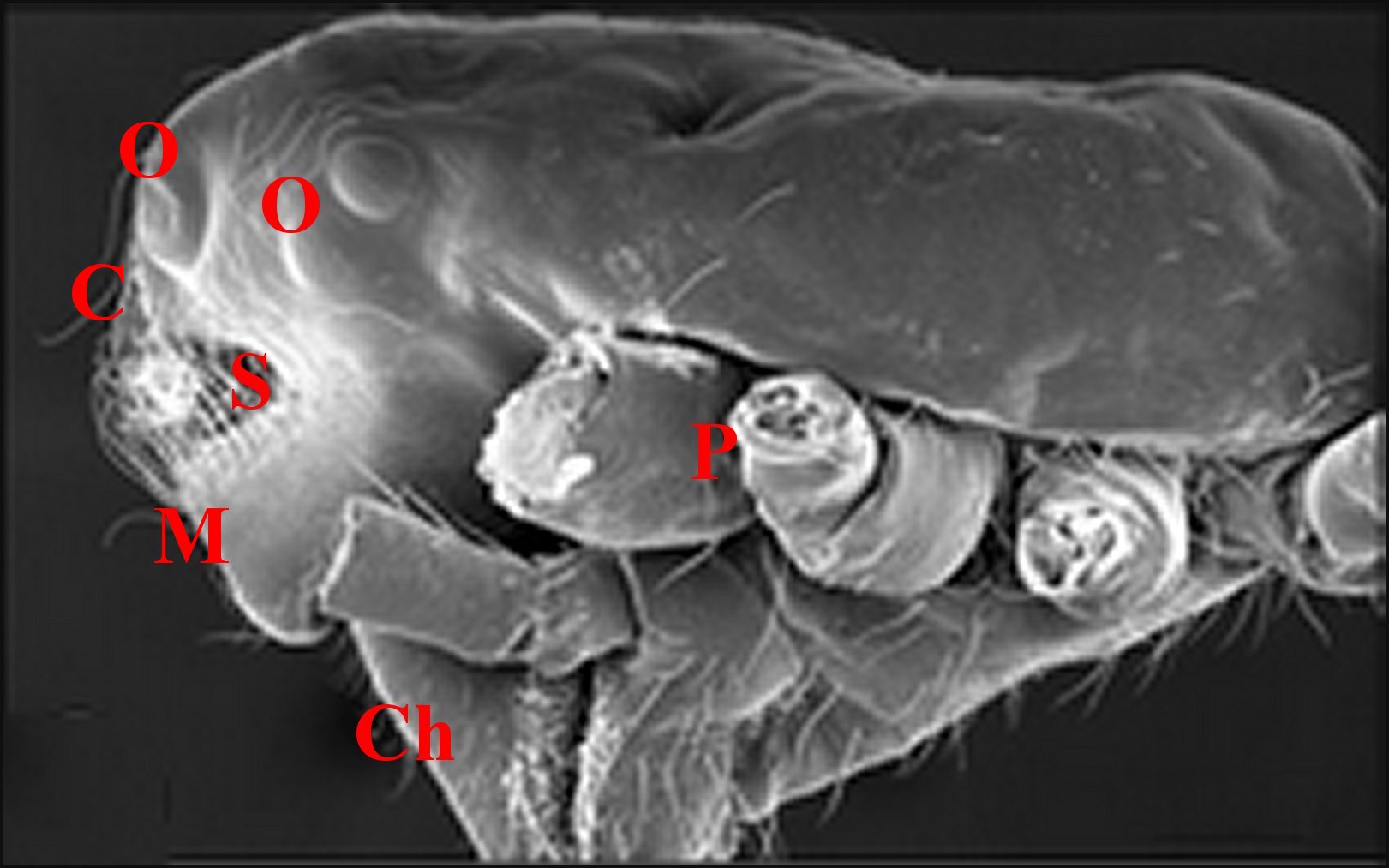
| Argyrodes
mâles : Bf, bosse frontale - E et flèches,
échancrure - H, poils - O, oeil - P,
pédipalpe -
Po, protubérance oculaire (© A.Lopez M.E.B.) |
|
 |
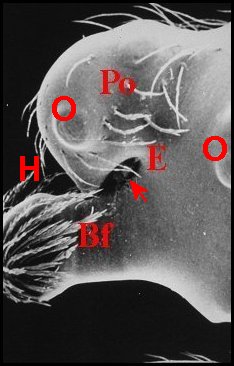 |
| Fig. 8 - Argyrodes nephilae , vue latérale gauche |
Fig.9 - A. rostratus nephilae,
idem. |
| Argyrodes mâles : Bf, bosse frontale - E
et
flèches, échancrure - H, poils - O,
oeil - Po,
protubérance oculaire (© A.Lopez M.E.B.) |
|
►Le
type 3 dit
nasuté paraît devoir
être réservé à Argyrodes nasutus
Pick.Cambr., de
Ceylan, dont la bosse frontale
se
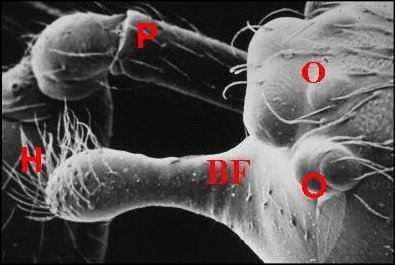
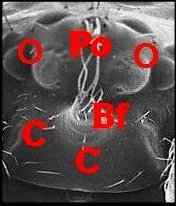
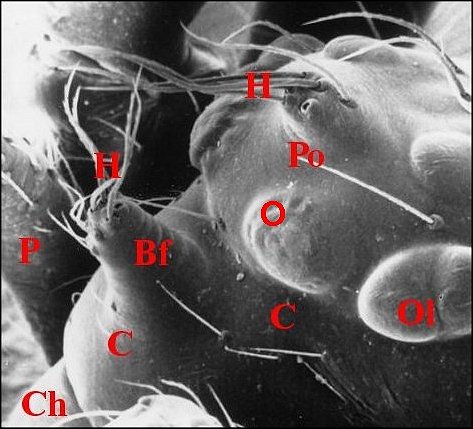
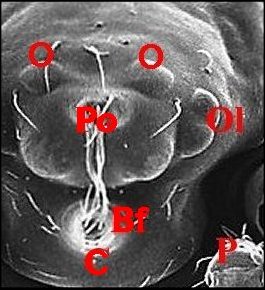
dispose en une sorte de “ nez ” caricatural (Fig.10 :
Bf ) ainsi qu'à Argyrodes
borbonicus et A.chounguii
Lopez,
nouvelles espèce, respectivement de la Réunion (Fig.14
annexe) où l'auteur l'a découverte en Aout 1986 (Fig.11
à 14) et de Mayotte
.https://fr.wikipedia.org/wiki/Argyrodes_chounguii (fig.2)

 |
 |
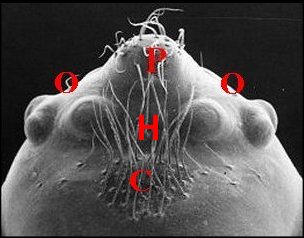
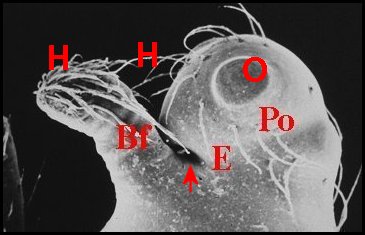
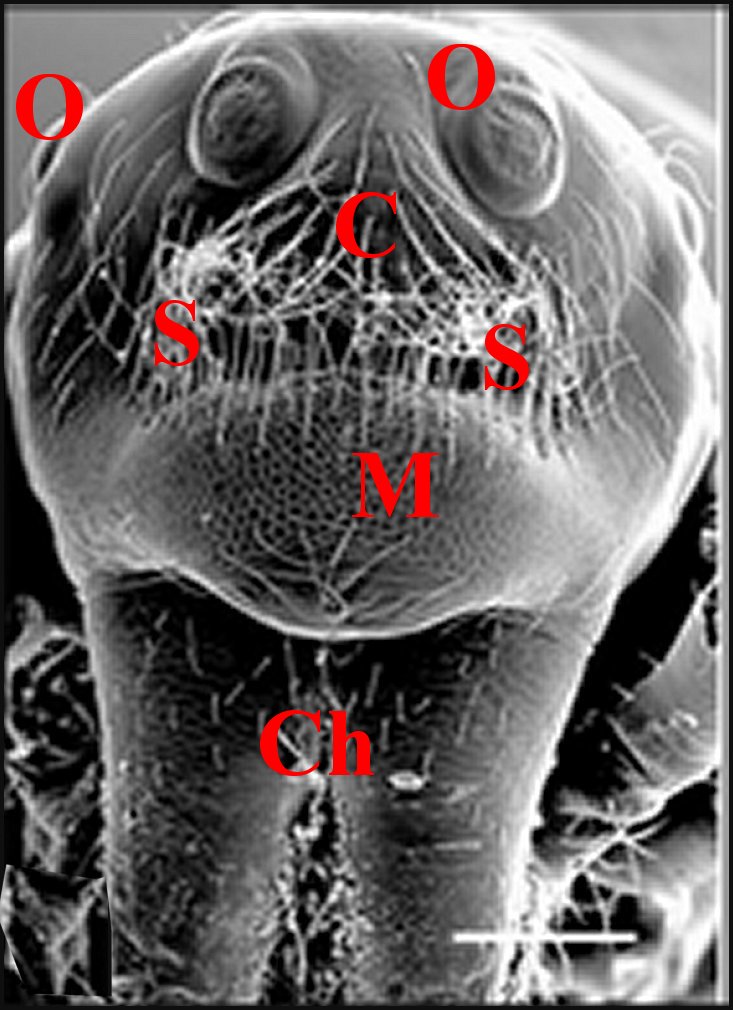
 |
| Fig.12 -Même Argyrodes, vue frontale | Fig.13 -Le même, vue latéro-supérieure gauche | Fig.14-
Le même, vue
supérieure ou dorsale |
| Argyrodes borbonicus : Bf,
bosse frontale - C, clypéus - Ch, chélicère - H,
touffe de poils - O,
yeux - Ol, yeux latéraux - P, pédipalpe -
Po,
protubérance oculaire (© A.Lopez M.E.B.) |
||
 |
|
Fig.14
annexe Argyrodes
borbonicus Lopez, mâle, vue latérale gauche
entière
D'après
Brice Derepas
|
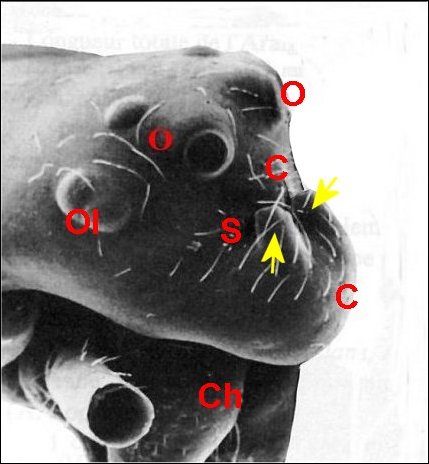
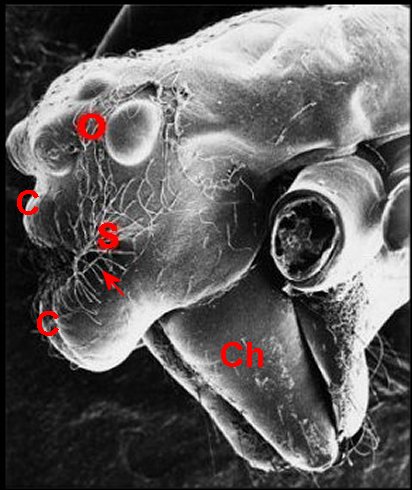
►Le type lippu
(Fig. 14,15) est
caractérisé par
une forte “bosse clypéale"
surplombant les chélicères
comme
une lèvre
inférieure éversée. Cette saillie forme la berge
antérieure d'un sillon
(S)
que garnissent des poils. Il
se rencontre
chez A.
amplifrons ( Fig.
15, 23) et Argyrodes ululans
(Fig.16), de Guyane française.
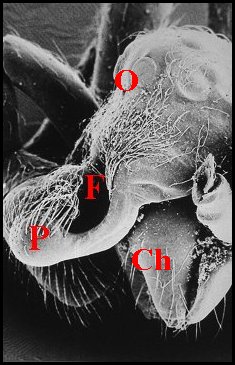
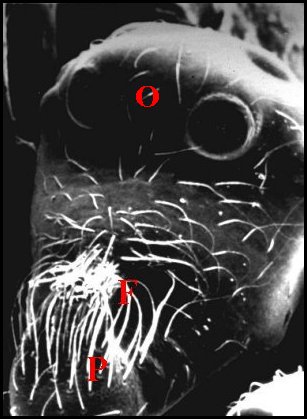
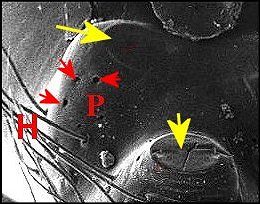
►Le type prognathe paraît
propre à
certaines autres espèces sud-américaines : Argyrodes
benedicti Lopez, également de Guyane
française (Fig.18,19), A.cochleaforma
(Fig.17), A. sullana, A.atopus,et
A. proboscifer.,
schématisées par Exline
& Levi (1962) sans en rechercher,
faute d'histologie, la signification anatomique profonde
(Note 1).
Le
clypeus saille
très fortement au-dessus des tiges
chélicériennes
(Ch), tel une mandibule,
cette protubérance (P), plus ou moins
incurvée, étant creusée d’une fossette poilue (F)
et pouvant avoir un aspect en “cuillère” (Fig.17)
 |
 |
 |
|
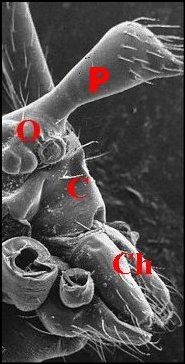
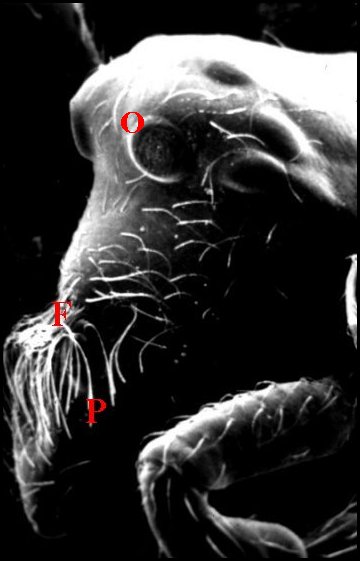
Fig.17 - Argyrodes cochleaforma ,
vue
antéro-latérale gauche |
Fig.18
- Argyrodes benedicti,
vue antérieure |
Fig.19
- Le même, vue latérale gauche |
|
Argyrodes mâles : Ch, chélicère -
F,
fossette - O, yeux - P, palpe - Pr, protubérance en
"mandibule". Les flèches soulignent les fossettes (© A.Lopez M.E.B.)
|
||
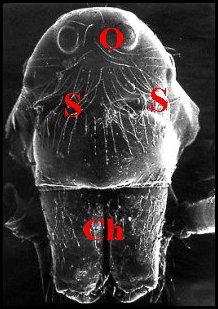
►le type camard
(par exemple Argyrodes
caudatus (Fig.20,21)
et A.cancellatus, tous
deux des Antilles françaises, est
caractérisé par l’absence de projections ; il montre un
clypeus (C) peu saillant,
haut, presque droit et parait donc aplati, comme
“écrasé”, si
on le compare aux types précédents ;
l’échancrure est
remplacée par deux sillons
transversaux
symétriques (S) étroits, presque “fermés” et
n’apparaissant bien que lorsqu’on
examine l’animal de face ou presque.
De plus,
l’étude au M.E.B. (Lopez,1979) montre
l’aspect du tégument
(lisse ou garni de reliefs en
“écailles”) et l’existence de saillies
accessoires plus ou moins tronconiques (Fig.22,24), en
cônes bas
tronqués dans le cas de l'espèce princeps cosmopolite Argyrodes
argyrodes (Fig.22,24) ou en "chicots" chez Argyrodes
amplifrons de Guyane (Fig.15, 23),
tendant à fermer l'échancrure
ou
le sillon.
Elle
met aussi en évidence la
structure des poils
(sillons, spinules…) et
surtout, ce qui est le plus important, l’existence d’orifices (Fig.24,25).
 |
 |
|
Fig.24
- Argyrodes argyrodes,
dessus de la bosse frontale
|
|
|
Argyrodes
mâles : H, poils - P, groupes de pores.
Flèches jaunes, saillies
accessoires, et rouges, pores
(© A.Lopez M.E.B.) Fig. 24 : en fait, Rhomphaea projiciens |
|
Ces
derniers
sont béants, sans bourrelet complet, parfois circonscrits
par deux lèvres, non
surélevés, le plus souvent de
niveau avec la surface tégumentaire, rarement dans le fond
de fentes étroites en “ boutonnières ”(Argyrodes
fissifrontella,
Seychelles) . Ils peuvent
être dispersés
sans ordre apparent (type acuminé) ou
réunis en deux
groupes
de 2 à 5 orifices
chacun, symétriques par rapport au plan sagittal (type
rostré). Dans
tous les cas, ils sont nettement séparés des poils, y
compris chez
l’espèce néozélandaise Argyrodes antipodiana
.Ils ne s’ouvrent pas
dans leur alvéole
basale, bien que Whitehouse (1987)
affirme le contraire par manque
d’étude
histologique ("Male
Argyrodes antipodiana secrete
a substance onto cephalothoracic protrusions ("knobs").The substance
apparently seeps out at the bases of hairs situated on
the knobs, then proceeds to cover
the hairs"). (Note
1)
Ils sont les pores excréteurs d’un organe glandulaire sous-jacent remarquable, déjà perceptible par transparence dans des céphalothorax éclaircis artificiellement (Fig.25).
|
|
Fig.25 - Céphalothorax d'un Argyrodes argyrodes mâle (Tunisie) éclairci au salicylate de méthyle.Bf, bosse
frontale
avec ses poils - E, échancrure - G, glande acronale
vue par
transparence
|
|
Sa
connaissance est fondée sur les études histologiques et
surtout sur les recherches électronomicroscopiques (Lopez,
1980a : Argyrodes argyrodes de type
“rostré” ; Lopez,1980b : Argyrodes cognatus, de
type
“acuminé") effectuées
essentiellement au Laboratoire souterrain du CNRS (Note
2).
Elles
en montrent respectivement la
structure d’ensemble et la structure fine de l’unité glandulaire.
3.1- Structure
histologique
Dans les
types
“rostré”
(Argyrodes argyrodes, A. zonatus,
A.elevatus,
A.nephilae)
(Fig.26 à 29), "lippu" (Argyrodes
ululans) (Fig.30)
et “camard” (Argyrodes
caudatus, A.cancellatus)(Fig.31),
un canal collecteur principal
aboutit à chacun des pores.
Il
résulte de la
confluence de canalicules
excréteurs de 2eme
ordre ; ces derniers sont eux-mêmes
formés par l’union de canalicules
excréteurs de 1er
ordre, chacun drainant
une cellule
sécrétrice ou adénocyte
et constituant avec elle
l’unité
glandulaire . Les canaux
excréteurs de
tous ordres sont entourés par des cellules canalaires. L’
ensemble de tous ces canaux constitue un appareil cuticulaire
s’étendant des portions
réceptrices aux pores
de surface.
Les
adénocytes se groupent
par
5 ou 6 en un petit lobule
assez bien individualisé
(Fig.27) que l'auteur avait d’abord interprété
(à tort !) comme
des
acini à lumière virtuelle
rappelant ceux des glandes
salivaires gnathocoxales (Lopez,1974a).
L'ensemble des lobules
constitue l’un des
deux massifs compacts
incurvés (Fig.26)ou des cordons
sinueux
(Fig.32,33) dont
est formée la
glande (Lopez,1974a ;
Lopez,1977 ; Lopez,1979).
Chaque massif ou cordon d' adénocytes aboutit
donc séparément à l’un des deux groupes de
pores
du sillon ou échancrure
par le système des canaux
interposés (glandes
“ arborescentes ”).
 |
 |
 |
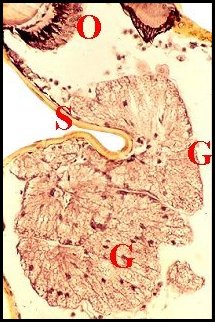
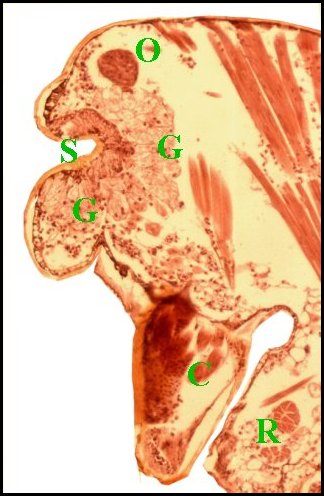
| Fig.26 - Argyrodes zonatus | Fig.27
- Argyrodes
zonatus,
coupe voisine. Détail |
Fig.28 - Argyrodes elevatus |
|
Glandes
acronales d' Argyrodes mâles, coupes
parasagittales du prosoma.
C,
chélicère - G,
massif
glandulaire incurvé - M, gnathocoxe - O, oeil - S,
échancrure clypéale
(© A.Lopez C.H.)
|
||
 |
 |
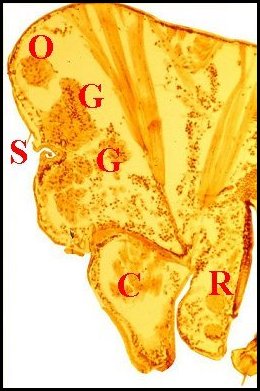 |
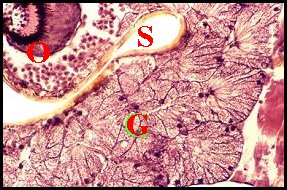
| Fig.29 - Argyrodes nephilae |
||
|
Glandes
acronales d' Argyrodes mâles, coupes
sagittales et parasagittales du prosoma.
C :
chélicère - G, massif glandulaire compact ou en cordons -
M, gnathocoxe - O, oeil - R, rostre - S, échancrure ou sillon
du clypéus (©
A.Lopez C.H.)
|
||
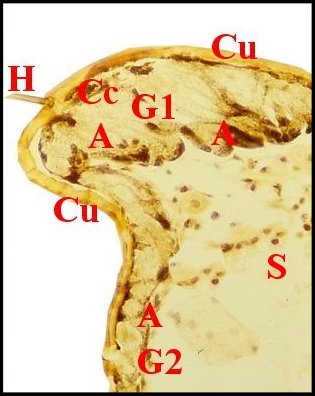
 |
 |
|
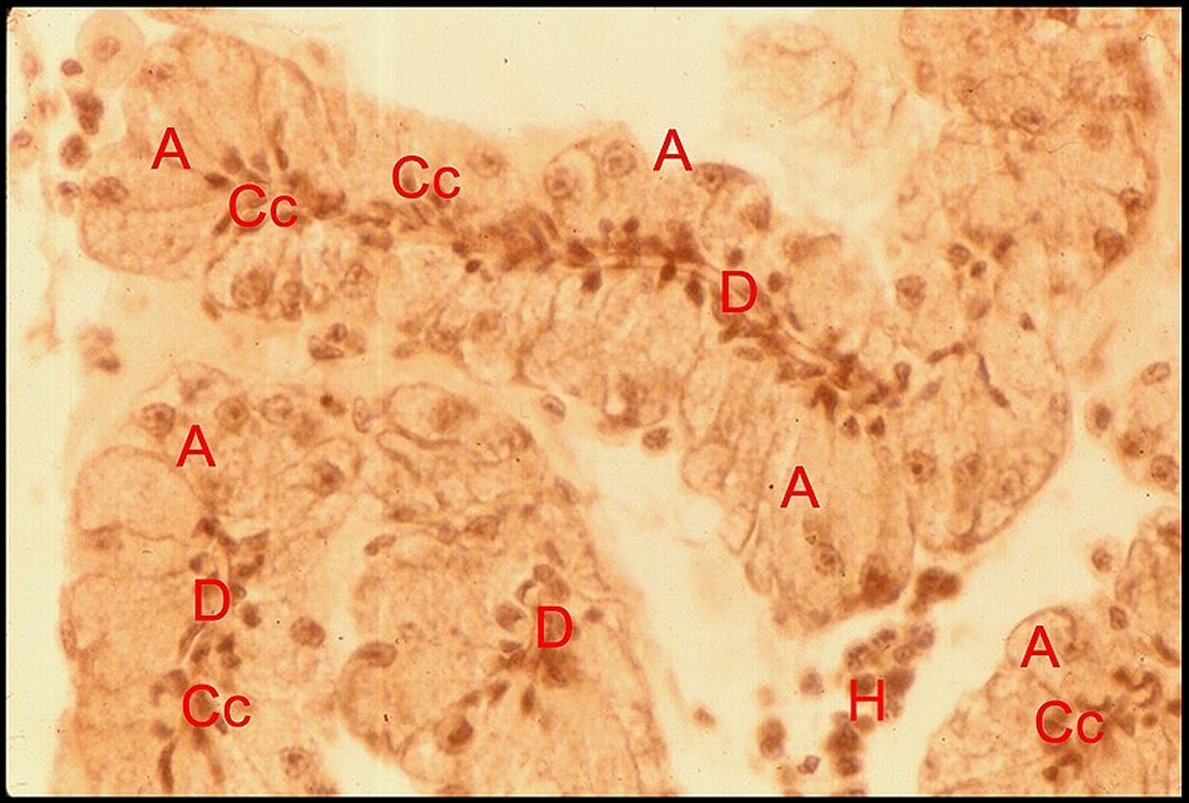
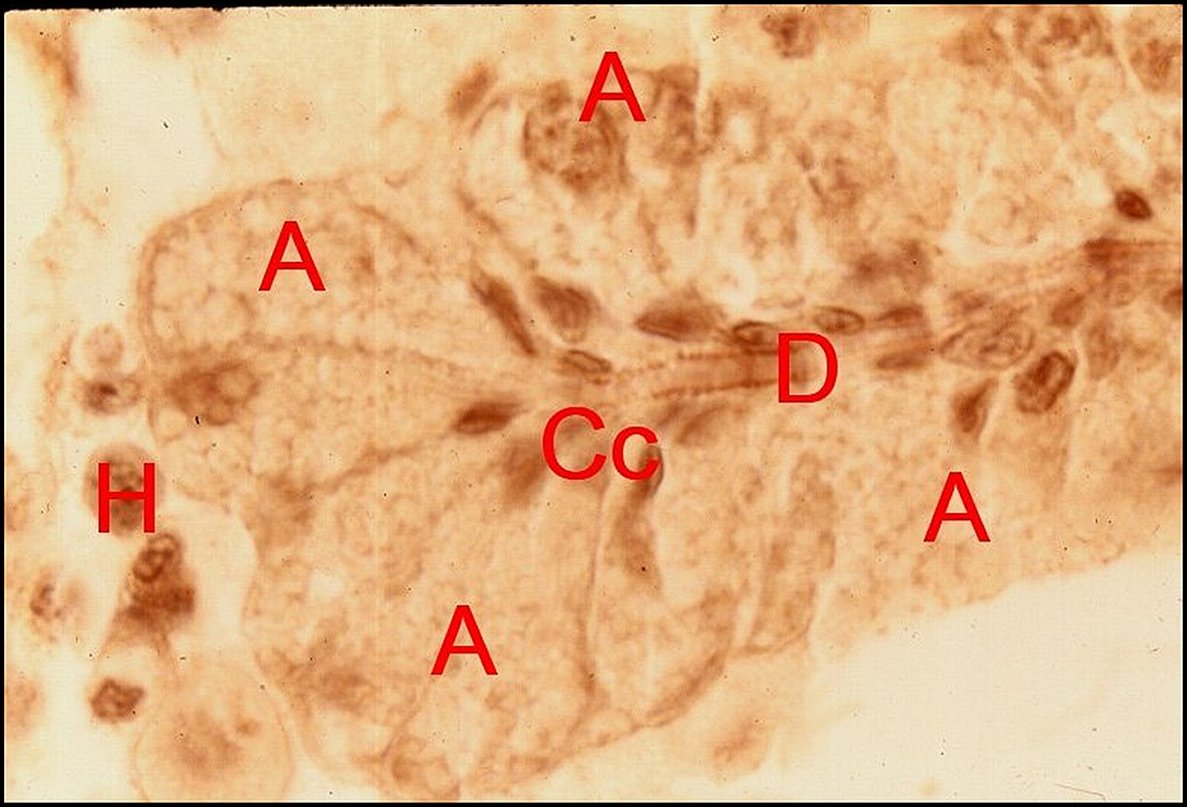
Glande
acronale d' Argyrodes caudatus mâle.
A,adénocytes - Cc, cellules canaliculaires - C, canal
excréteur d'un cordon.
H,
hémocytes (©
A.Lopez C.H.)
|
|
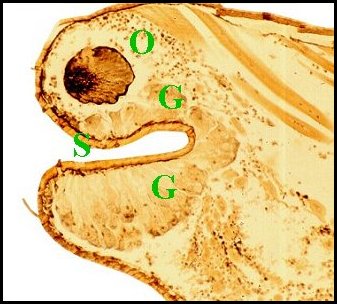
Toutefois, chez l’espèce Argyrodes fissifrontella (Seychelles),
de morphotype rostré, l’ensemble
de l’organe ne paraît pas bilobé mais impair (Fig.34,35) ;
ses
unités glandulaires
sont isolées ou en lobules
lâches ; leurs canaux
ne forment pas un système ramifié mais restent
indépendants comme dans le type
suivant (“acuminé”) ; de plus,
leurs terminaisons
ne se limitent pas à la bosse
frontale mais s’ouvrent
également
sur la protubérance oculaire
(Fig.35) (Lopez,1981).
Dans
le type
“acuminé” (Argyrodes
cognatus) un massif
glandulaire également impair occupe la protubérance
sus-oculaire en "tourelle"
(“turret gland”) et se prolonge en “languette” sous le tégument
clypéal (Fig.36, 37).Il est formé par
des
unités
juxtaposées
bien séparées ou à disposition lobulaire peu
marquée, chacune d’elles comprenant un adénocyte, une
seule cellule canalaire et un
canal excréteur de 1er
ordre, aboutissant
directement à un
pore, parfois après
s’être
fusionné avec un voisin juste avant cette terminaison. Comme
chez
A.fissifrontella, il
n’existe pas de système
excréteur ramifié
(glandes “simples”).
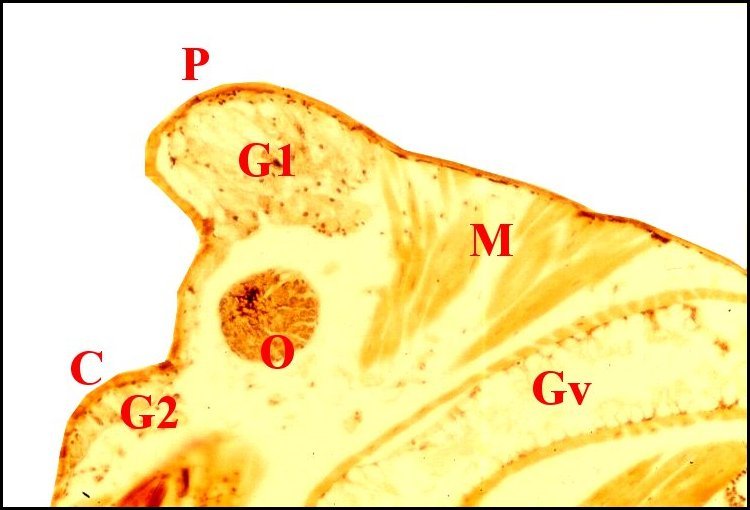 |
 |
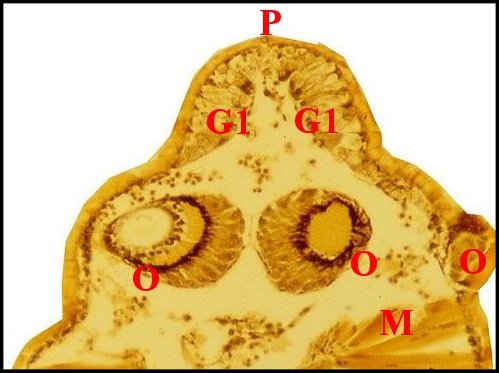 |
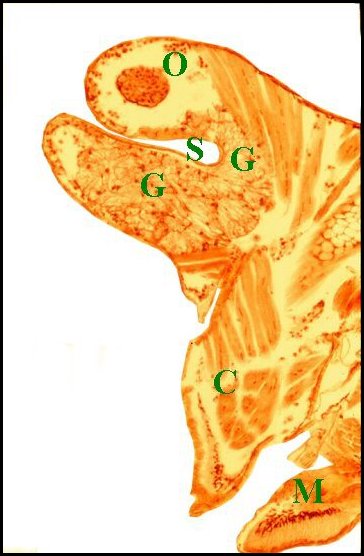
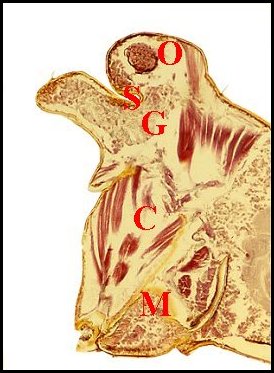
| Fig.36
- Argyrodes cognatus, coupe sagittale |
Fig.37
- Argyrodes cognatus,
coupe sagittale, détail |
Fig.38- Argyrodes cognatus,
coupe transversale |
|
Glande
acronale d' Argyrodes cognatus mâle
en coupes sagittales et transversale. A,
adénocytes des unités - C, clypéus - Cu, cuticule
- G1, massif glandulaire de la "tourelle" - G2, languette
clypéale - Gv, glande à venin - M, muscle - O, oeil
- P, protubérance sus-oculaire ("tourelle") -
S,
sinus sanguin (©
A.Lopez C.H.)
|
||
3.2- Ultrastructure
3.2.1- Unité glandulaire
Elle
représente une structure anatomo-fonctionnelle de base,
autonome, bien
individualisée, constituée par une cellule glandulaire
ou adénocyte et
son canalicule
excréteur de 1er ordre ou canalicule conducteur
(Schéma).
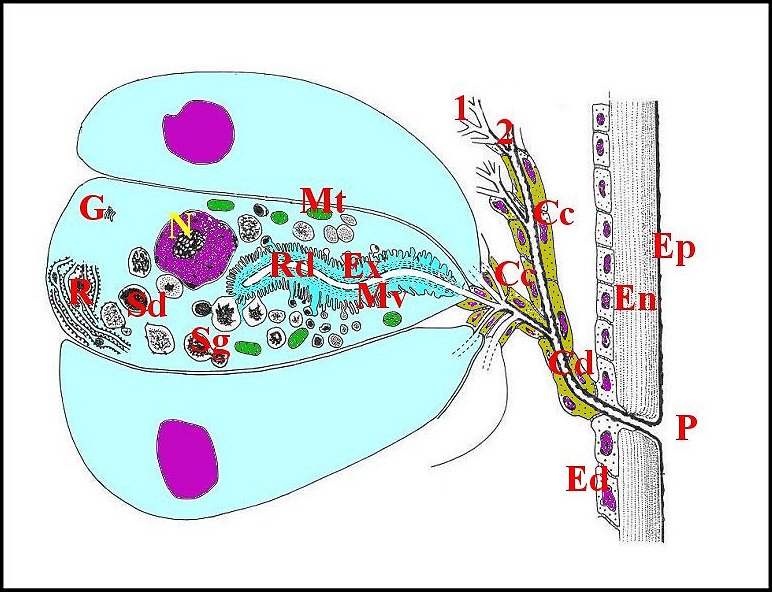 |
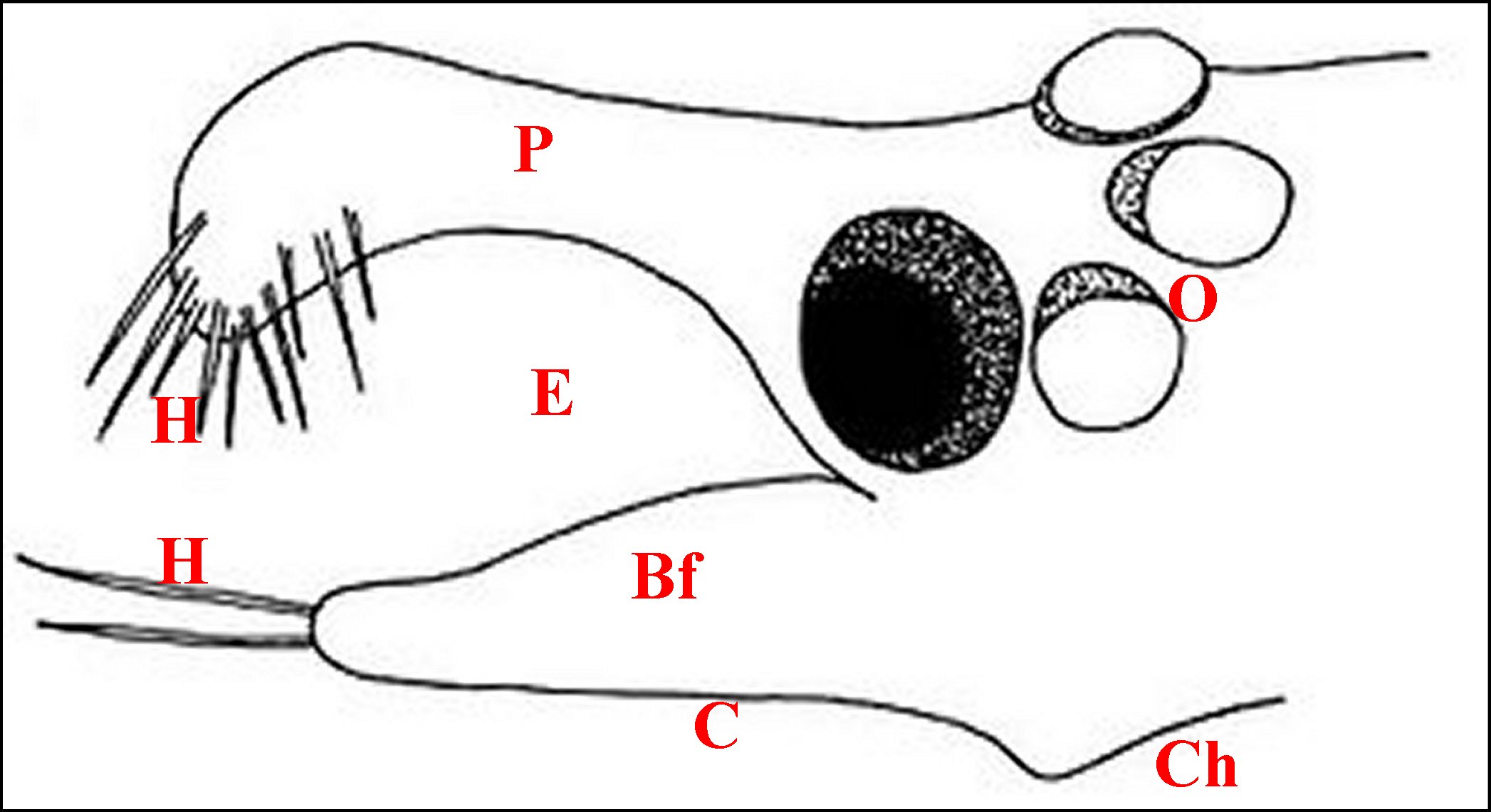
| Schéma
- Unité
glandulaire d'Argyrodes argyrodes
que flanquent
deux unités voisines et la totalité de son appareil cuticulaire |
| Cc,
cellules canalaires - Cd, canal excréteur principal - Ed,
épiderme - Ep, épicuticule - En, endocuticule - Ex,
cavité extracellulaire - G, golgi - Mt, mitochondrie - Mv,
microvilli - R, réticulum granulaire - Rd, canalicule
récepteur - Sd, sécrétion dense - Sg,
sécrétion granuleuse - 1 et 2, canalicules
excréteurs de 1er et 2eme ordre. |
333-2-1a Adénocyte ou
cellule glandulaire
L’adénocyte
est pyramidal ou
piriforme, long en
moyenne de 65 µm, pourvu d’un noyau
vésiculeux
généralement basal, n’excédant pas 9 µm, et
d’ un cytoplasme assez
compact et basophile en périphérie, d’aspect
vacuolisé
et spongieux dans la majeure partie de son étendue, sans
sécrétion figurée. Il renferme un canalicule axial
surtout
visible en coupe transversale, et qu’entoure une zone arrondie en
“réservoir” acidophile,
faiblement PAS +.
Il est enveloppé par une gaine de lame basale le
séparant du sinus
hémolymphatique adjacent.
Sa membrane plasmique
ne
présente pas de dispositifs (replis) en augmentant la surface au
niveau
du pôle externe
(basal), qui
est régulièrement arrondi, et des faces latérales,
qui sont à peu
près planes. En revanche, elle pénêtre trés
profondément dans le pôle
apical qui présente ainsi
une invagination de l'espace
extracellulaire en "cul
de sac ” ou
“doigt de gant”. Ce “réservoir”
occupe en longueur la
moitié de l’adénocyte,
est sinueux, bordé par des microvillosités
(considérées d’abord
à tort comme des canaux intra-cytoplasmiques ! : Lopez,1975) et
renferme un canalicule
récepteur en position
axiale (Schéma, Fig.41,42,43). Les
microvillosités (Fig.37
à 40) sont courtes et espacées près de
l’apex, plus longues, plus
nombreuses et resserrées dans sa partie distale,
présentent des flexuosités et sont soutenues par du
cytosquelette (quelques microtubules,
surtout des filaments d’actine
réunis en faisceaux qui
s'ancrent sur une densification
apicale).
Le
noyau est
plus ou moins déformé par les grains
de sécrétion, renferme un nucléole volumineux et
une chromatine
finement grenue, dispersée, ou réunie en petites mottes
éparses.
Les organites
subcellulaires sont
des mitochondries
ovoïdes dispersées
dans le hyaloplasme, des ribosomes
remarquablement nombreux et surtout, un réticulum
endoplasmique extrêmement
développé et de type rugueux. Il se
présente sous la forme de tubules
irréguliers,
surtout localisés en périphérie, au voisinage du
plasmalemme, et de
citernes,
très variables dans leur aspect et leurs dimensions, occupant
surtout le reste de la cellule.
Les empilements membranaires (“dictyosomes”)
qui
caractérisent en général un appareil
de Golgi typique paraissent absents.
En
revanche, une sécrétion
abondante est
élaborée dans certains adénocytes.
Elle se
présente sous la
forme
de grains (Fig.37,39)
contenus dans des vésicules
semblant
issues du réticulum
qui les
entoure : les unes renferment un matériel peu dense, floculant,
irrégulier et grossissent par confluence ; les autres, non
coalescentes, ont un contenu plus opaque, homogène et
subsphérique
(Fig.37). Les mêmes vésicules
peuvent fusionner et extrudent leur contenu entre les bases des
microvillosités. Cette
substance, finement grenue, pénètre ainsi dans l' espace
extracellulaire, autour du canalicule
récepteur (Schéma).
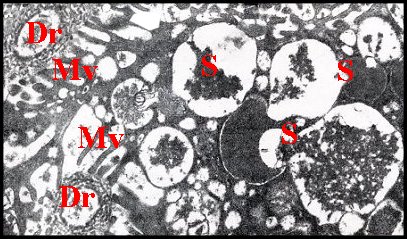 |
| Fig.39 - Adénocyte de glande acronale et son canalicule récepteur. |
| Glande
acronale d' Argyrodes argyrodes mâle Dr, canalicule récepteur (deux sections) - M, mitochondrie - Mv, microvilli - S, sécrétion en grains (© A.Lopez M.E.T.) |
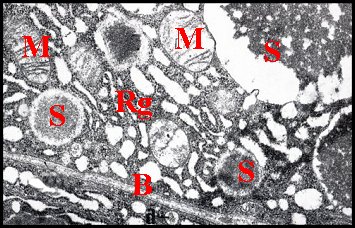 |
|
Fig.40 - Adénocyte de
glande acronale : organites.
|
|
Glande
acronale d' Argyrodes
argyrodes mâle
B, basale - M, mitochondries - Rg, réticulum granuleux - S, grains de sécrétion (© A.Lopez M.E.T.) |
3.2.1.b-Canalicule récepteur (portion réceptrice)
Il présente une lumière de 0,5 µ de diamètre et une paroi (épaisseur : 0,2 à 0,3 µ) comprenant deux couches : couche interne entourant la lumière, relativement homogène, perforée, et couche externe plus épaisse, lâche, d’aspect vermiculé, formée par des strates discontinues grossièrement concentriques se disposant en “ filtre ”(sieve) (Fig.39,40). Le matériel sécrétoire finement grenu ayant gagné l' espace extracellulaire se retrouve dans les interstices de ce filtre, puis rejoint la lumière par les pores de la couche interne et s’y accumule.
Le canalicule récepteur entouré par les microvillosités correspond à un “appareil terminal” (“end apparatus”) typique (Schéma, Fig.39). Il s’abouche à la portion conductrice qui est dans son prolongement direct.
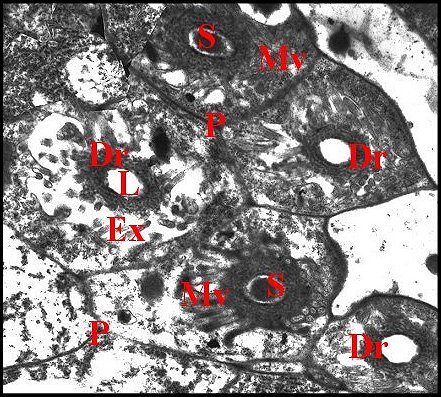
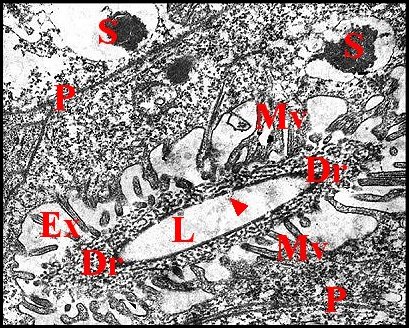
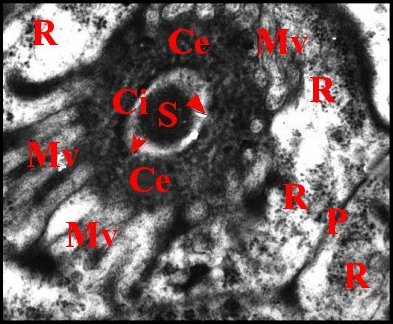
3.2.1.c-Portion
conductrice ou excrétrice
Elle
est formée par des canalicules
et
leurs propres cellules dites "canalaires" ou "enveloppes".
Un canalicule
excréteur ou conducteur
peut être soit simple, représentant sa
totalité lorsqu’il demeure isolé jusqu’à la
terminaison (glande également “simple”)(Fig.34,35), soit de 1er
ordre
lorsqu’il
conflue ensuite
(glandes “ramifiée”), dans les deux cas d’un même
diamètre, subcylindrique et à paroi (épaisseur 600
Å) réduite à une seule couche, homogène,
régulière, formée par de l’épicuticule.
Les
canalicules
d’ordres supérieurs (2eme ordre et excréteur
principal)
ont un diamètre plus grand
(1,2 µ et 2 à 5µ), une lumière plus large
et une paroi formée
cette fois par 2 couches :
l’une interne,
épicuticulaire,
dense, opaque et
homogène comme
celle du canal de
1er ordre,
mais d’épaisseur irrégulière, paraissant
festonnée en coupes transversales ; l’autre externe,
dérivée de l’exocuticule
(?),
plus claire, striée, montrant des fibres
hélicoïdales
et concentriques.
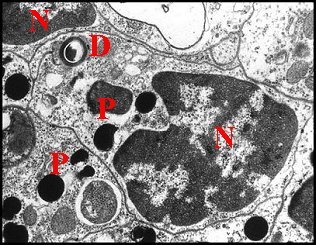 |
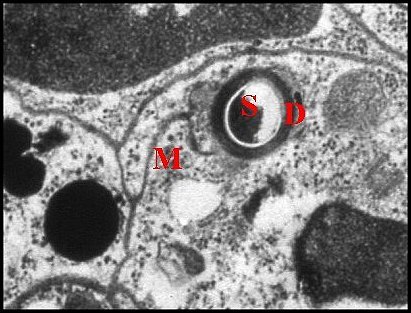 |
| Fig.44 - Canal excréteur et sa cellule canalaire | Fig.45 -
Les mêmes, détail |
|
Argyrodes cognatus
:
canal excréteur simple et sa cellule canalaire
D,
canalicule - M, méso - N, noyau ; P, pigment ; S,
sécrétion (© A.Lopez M.E.T.)
|
|
En ce qui concerne les cellules
canalaires (Schéma), il n'y en a
qu' une seule lorsque le
canal est “simple” (Argyrodes
cognatus)(Fig.44,45), plusieurs qui
se
succèdent en gaine continue lorsque la glande est
ramifiée (Argyrodes
argyrodes), dont
une proximale, autour du canalicule de 2eme ordre, et les autres
distales,
autour du
conduit principal. Dans tous les cas l’adénocyte
s’unit
à la cellule
canalaire adjacente par une jonction
adhérente annulaire (zonula
adherens). Peu visibles dans les
coupes
histologiques (pigmentation
mélanique
éventuelle, long noyau
très basophile), les cellules
canalaires montrent en M.E.T.,
qu’elles s’enroulent autour des
conduits et renferment dans leur hyaloplasme
clair des microtubules
ainsi
que
quelques grains de pigment
mélanique. Le noyau
est
très allongé, aplati, indenté, réniforme en
coupe transversale et
contient
une chromatine
abondante, dense et marginée. L’enroulement de chaque cellule
détermine une invagination
en méso de son plasmalemme (Fig.45), avec jonction
septée. D’autres jonctions l’unissent
aux voisines.
4 -Commentaires
4- 1 Sur
le
plan anatomique, la
structure fine des unités constituant la
glande
acronale permet de les rattacher à la classe 3
des
cellules glandulaires
épidermiques d' Arthropodes dans sa forme la plus
simple
: un
canal est d’abord
entouré par une seule cellule
glandulaire
où il débute,
puis par une cellule canalaire
et
est enfin en continuité avec la cuticule au
niveau de sa terminaison. Noirot
et Quennedey (1974) ont défini
pour la première fois cette classe chez les Insectes où
existe
également
un “appareil terminal”,
sensé
caractéristique de glandes
exocrines tégumentaires
défensives ou
sécrétrices de phéromones.
On
sait par ailleurs que dans de
nombreuses glandes exocrines
d’Insectes (salivaires, dermiques,
annexes du tractus génital), l’appareil
de Golgi ne
paraît pas
polarisé, ne se dispose pas en dictyosomes
mais paraît
uniquement
formé par des vésicules.
Il en est probablement de
même
pour la glande acronale des Argyrodes
où les amas
vésiculaires marqueraient l’emplacement de réseaux
transgolgiens.
Les
unités peuvent
être discrètes et
isolées (types “acuminé” et “nasuté”)
ou former un organe glandulaire anatomiquement défini, plus ou
moins volumineux et massif (autres types) montrant
généralement une bipartition visible dans les coupes
histologiques transversales et la disposition symétrique de
pores au M.E.B.
La glande acronale d’Argyrodes
fissifrontella se
présente comme une variation importante pouvant exister aussi chez d’autres espèces, et
qui a valeur de stade
évolutif.
4- 2 Sur
le
plan
fonctionnel,
la glande acronale est étroitement liée au comportement
sexuel des Argyrodes.
Jusqu’aux travaux pionniers de l'auteur,
les
conformations étranges du prosoma
des mâles ont été considérées
comme “ des bizarreries de la Nature ….évidemment sans
utilité ” (Berland, 1932) et
illustrées par Millot
(1968 : fig.479, p. 696) sans tentative d’explication alors
qu'il
était un remarquable histologiste ! (Note 1)
Il
convenait de rechercher dans l’éthologie d’ Argyrodes
certains traits de comportement pouvant relier des
hypothèses fonctionnelles à l’existence de la glande.
La relative immunité dont
bénéficient les Argyrodes
dans leurs
rapports
avec l’Araignée-hôte n’est nullement concernée.
La
glande
acronale ne peut
être considérée comme un organe de défense
élaborant
une substance répulsive ou vulnérante : elle manque chez les femelles ; l’absence de musculature
compressive et la terminaison des canaux excréteurs dans
une région anfractueuse conformée en cul-de-sac
n'impliquent pas une projection de substance ni sa libération
massive vers un agresseur lors des "stress".
 |
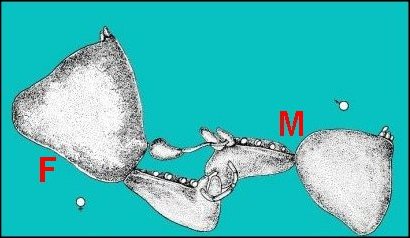 |
 |
|
Fig.46
- Argyrodes fissifrontella (Seychelles)
Préludes,
accouplement
(Ancienne diapo. de A.Lopez)
|
Fig.47.- Argyrodes zonatus, accouplement
: au centre, positions ; à droite, détail des
prosomas (schéma d'après Legendre,1960) |
|
| F,
femelle - M, mâle - P, palpe et son bulbe copulateur.
Flèche :
étreinte de la protubérance oculaire (?). |
||
 |
|
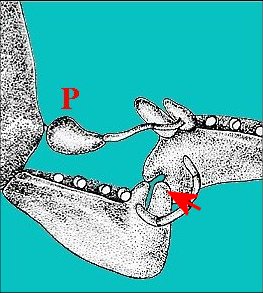
Fig.48.-
Argyrodes argyrodes (Cap Vert), accouplement
: étreinte du céphalothorax mâle par la femelle.
(d'après
B.Knoflach, Internet)
|
|
B, bosse frontale - C,
chélicères femelles - F, femelle - M, mâle - P,
protubérance oculaire .
|
 |
 |
|
Fig.49.-
Même couple
: insertion du palpe mâle droit dans l'épigyne de la
femelle
|
FFFFig.50.- Même couple : sécrétion vraisemblable du palpe mâle droit introduite dans l'épigyne de la femelle pour l'obturer |
|
E, épigyne - F, femelle - H,
haematodocha du palpe droit - M, mâle - S,
sécrétion palpaire pouvant se concréter en
obturateur de
l'épigyne
(d'après
B.Knoflach, Internet)
|
|
De plus, dans le cas du type “rostré”, le seul, semble-t-il, dont l’accouplement et ses préludes aient été étudiés (Legendre, 1960 ; Withehouse,1987) jusqu'à la rédaction initiale de ce texte puis ensuite photographiés avec talent mais sans commentaires anatomiques (Fig.48 à 50), les chélicères de la femelle étreignent la bosse frontale du mâle, leur “ doigt mobile pénétrant ” selon Legendre ”dans le repli” - plus précisément l’échancrure- “formé par la bosse frontale et le clypeus” -plutôt la protubérance oculaire. Il est probable qu’au cours de cette manœuvre durant laquelle “ le mâle est littéralement prisonnier des chélicères…”, les parties buccales de la femelle entrent en contact intime avec l’échancrure et la sécrétion clypéale qui doit s’y déverser. De futures recherches anatomiques axées cette fois sur le modelé des appendices péri-buccaux femelles, pourraient établir ultérieurement certaines correspondances entre ce dernier et les reliefs clypéaux mâles. Il ne semble pas que la sécrétion soit consommée par la femelle (contrairement à ce qui se passe chez certains Insectes comme les Dictyoptères) mais plutôt qu’elle s’évapore dans la dépression où les poils pourraient jouer un rôle pour la concentrer et (ou) la diffuser. Sa fonction est celle d’une phéromone qui facilite l’accouplement, donc aphrodisiaque, soit en inhibant momentanément l’agressivité de la femelle dont Legendre (1960) a d’ailleurs souligné une “certaine torpeur” (effet pacificateur), soit en provocant son excitation sexuelle (“arousal”, selon Withehouse,1987) ce qui paraît plus douteux.
En
outre, et contrairement à une
autre assertion de Whitehouse (1987),
rien ne prouve que la substance
soit élaborée en permanence par le mâle qu’elle
marquerait
ainsi chimiquement.
Notons
déjà que la copulation s’accompagne
de la mise en place d’un “bouchon d'
accouplement”
(“mating plug”) sur l’épigyne
femelle,
sécrétion
possible
du palpe
mâle intromis (Fig.50), entrant dans le cadre étrange
des obturateurs
génitaux présentés
ailleurs dans ce sous-site et que nous
ne redécrirons donc pas à
nouveau ici.
4-3.- Sur le plan
systématique, le
genre Argyrodes sensu stricto a été seul
envisagé jusqu'alors.
En fait, il appartiendrait à une sous-famille
plus ou moins bien délimitée et dont certains taxons,
d'ailleurs contestables, pourraient être remis plus tard en synonymie.
4-3-a - Les Argyrodinae Simon,1894 (Conopisthinae Archer, 1950).
Ils comportent, outre Argyrodes,
"chef de file", les genres Ariamnes et Rhomphaea, autrefois
et souvent encore rattachés encore au précédent, ainsi
que Neospintharus ,
Faiditus, Deelemanella
et
Spheropistha. A l'exception des deux derniers, ces
genres ont été
présentés par I.Agnarsson
(2004) , axée sur la seule anatomie externe, en soulignant
la morphologie étrange des mâles (Fig.51 à 54) mais
sans
prospection histologique (Note 1) et bien
évidemment, sans la moindre allusion
à une glande acronale
ou clypéale
sous-jacente pouvant
l'explique. Par ailleurs, elle signale des "filières
glandulaires épiandriques"("epiandrous glands
spigots") et des "médiators stridulatoires abdominaux sur
crête parallèle avec le pédicule" ("abdominal
stridulatory picks on ridge, parallel with pedicel"moins sous silence,
dans sa bibliographie, l'appareil
épigastrique et les sensilles
pédicellaires.
 |
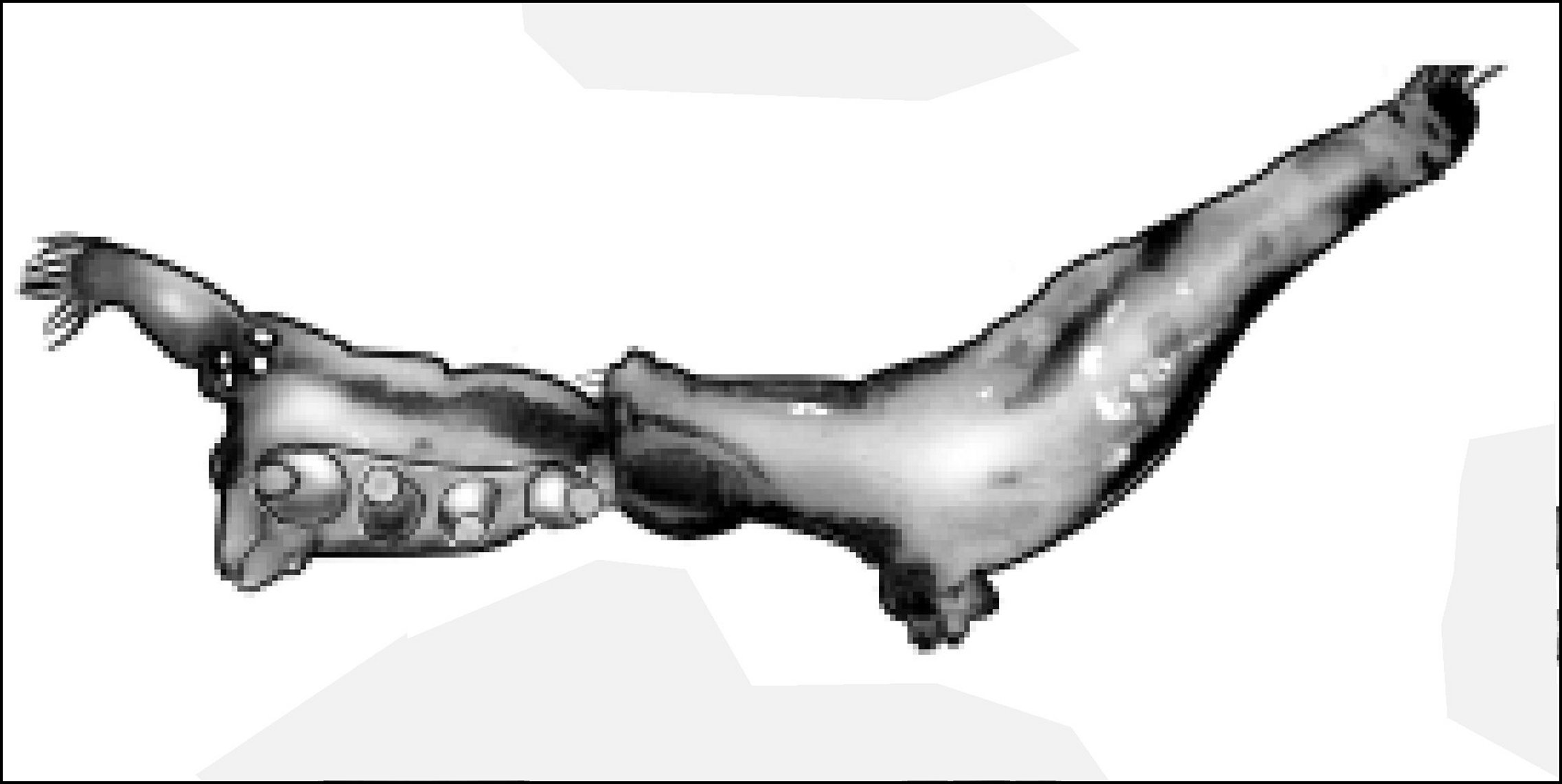 |
 |
 |
|
| Fig.51- Ariamnes |
Fig.52- Rhomphaea |
Fig.53 - Faiditus |
Fig.54 -Neospintharus |
|
|
Morphologie
et habitus des mâles d' Argyrodinae
autres qu' Argyrodes sensu stricto
d'après Agnarsson (2004 : fig.94)
|
|
|||
Le genre Ariamnes,
créé par Thorell en 1869, réunit des
Araignées Theridiidae
grêles (Fig.51,55,56), à prosoma
étroit et abdomen de couleur variable remarquablement allongé,
cylindrique, trés flexible, vermiforme ou bacilliforme,
d'où les noms spécifiques suggestifs qui ont
été créés (colubrinus,
columnaceus,
cylindrogaster,flagellum,
longissimus...),
évoquant des "fouets"
ou des "rameaux" pour les anglophones ("whip" ou "twig" spiders) et se rapprochant par son
aspect des Phasmes ou Bacilles, dans la classe des Insectes http://faune-flore-languedocienne.alwaysdata.net/garrigue/Insectes_Cheleutopteres.html
Les parties clypéale et
oculaire du prosoma
mâle peuvent avoir un aspect banal (Fig.55 à 57), proche
de celui de la femelle, montrant tout au plus, de profil et, entre les
yeux, une discrète intumescence garnie de poils (Fig.51,57). Toutefois, chez
certains taxons des îles Hawaï à noms spécifiques vernaculaires surprenants (Fig.58,59), il existe une bosse frontale et une protubérance interoculaire,
pileuses, trés saillantes en avant et séparées par
une échancrure, ce qui
les rattache au morphotype rostré des Argyrodes
sensu stricto. Une glande acronale
est, sans nul doute, présente dans tous les cas,
même à minima et devrait être confirmée
par l'histologie et le M.E.B.
 |
 |
 |
 |
 |
| Fig.55 - Ariamnes
colubrinus, mâle.
Vue latérale droite D'après H.Vanuytven |
Fig.56
- Ariamnes
sp., mâle,
Tasmanie. Vue dorsale du prosoma |
Fig.57 - Ariamnes,
mâle D'après Agnarsson (schéma) |
Fig.58 -Ariamnes kahili,
mâle Hawaï D'après Gillespie et Rivera (schéma) |
Fig.59 - Ariamnes
melekalikimaka, mâle Hawaï d'après Gillespie et Rivera |
|
Bf,
bosse frontale -C, clypeus - Ch, chélicère ou son
emplacement - E, échancrure - H, poils -
O, yeux - P, protubérance interoculaire. |
||||
Rhomphaea
est un genre riche en
espèces, surtout exotiques, telles R.metaltissima
de Bolivie, du Brésil (Fig.62), et surtout que R.(ex Argyrodes)projiciens (Fig.5,24) répartie en
Amérique des USA à l'Argentine et introduite
en Inde. Le même groupe est représenté en France
par deux
espèces découvertes lors de battages
végétaux : Rhomphaea nasica (Simon,1873) (Fig.61) de
Corse,
Port-Cros, d'Occitanie
(Maurel,2014,Villespassans : Hérault
: une femelle seulement), région où demeure Lopez André,
et Rhomphaea
rostrata (Fig.60)
citée à tort comme une Argyrodes
par
Emerit (2002) (Note
3)qui l'a
récoltée dans l' Hérault,
en Dordogne et toujours accidentellement. A l'exception, trés
curieuse, de
celui de cette dernière ("Male prosoma 1-1.5 mm long
without tubercle" (Fig.60), "le mâle ne porte aucune
protubérance frontale contrairement au curieux R.nasuta de Corse", Berland, 1926), les
mâles (Fig.61,62) sont pourvus d'une protubérance
interoculaire évoquant la "tourelle" d' Argyrodes
cognatus (Fig.3), ce qui
permet de lier
leur prosoma au morphotype acuminé
La présence d'une glande acronale y est
assurée grace à
l'observation en fort grossissement de ses pores excréteurs
(Fig.24)
, discernables chez deux autres espèces (Fig.61,62)
où ils s'associent à des poils apicaux en "toupet ".
 |
 |
 |
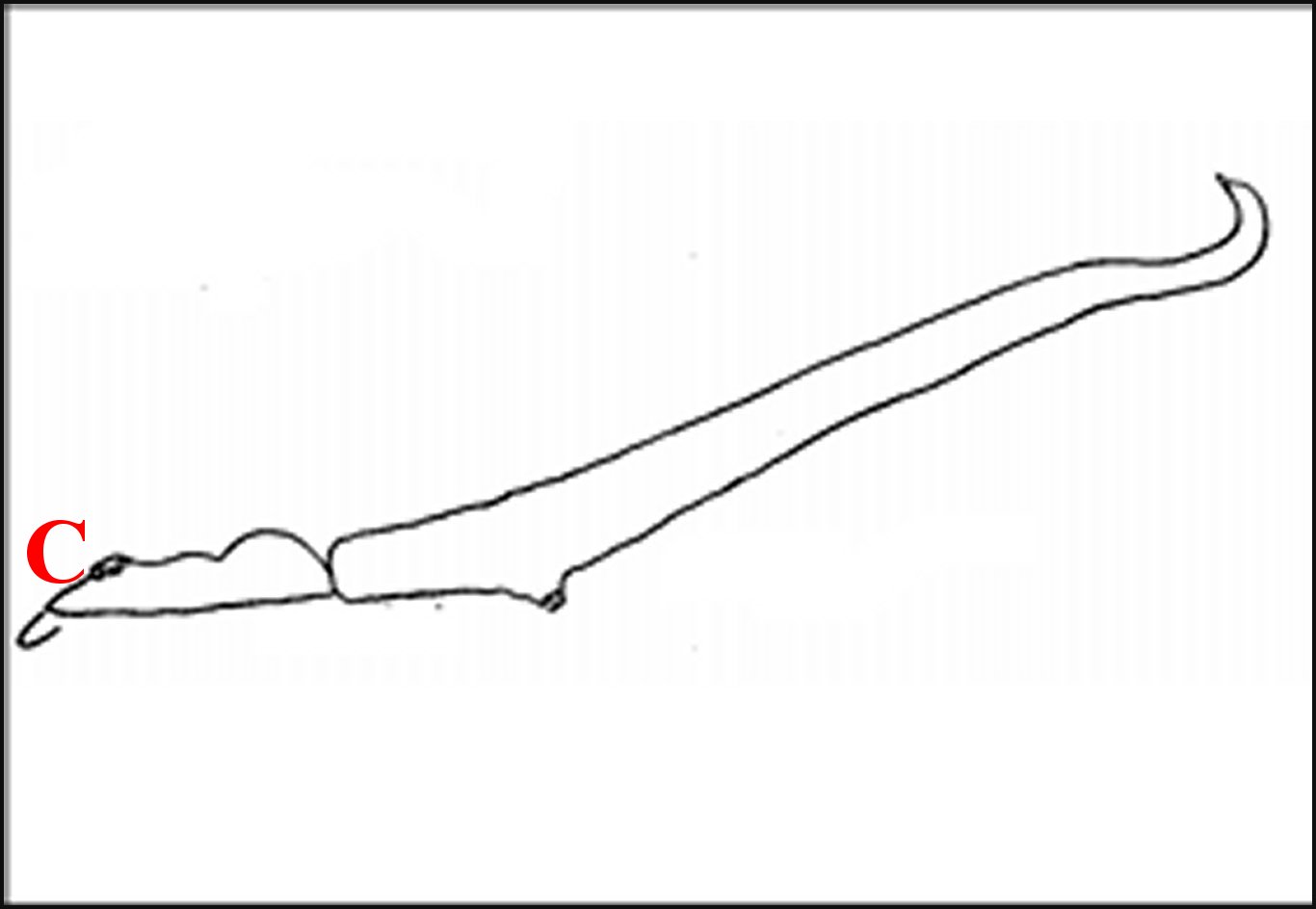
| Fig.60
- Romphaea
rostrata , mâle, de profil gauche, sans les pattes.
Prosoma, à gauche, à clypeus (C) trés oblique,
dépourvu de tubercule D'après Wunderlich,1987 (esquisse) |
Fig.61
- Romphaea nasica,
mâle, de profil gauche, sans les pattes. Prosoma pourvu
d'un tubercule interoculaire procurvé D'après Oger-J.P.Taberlet (photo) |
Fig.62 - Romphaea
metaltissima, mâle,
de
profil gauche, sans les pattes. Prosoma pourvu d'un tubercule
interoculaire
D'après Agnarsson (M.E.B.) |
|
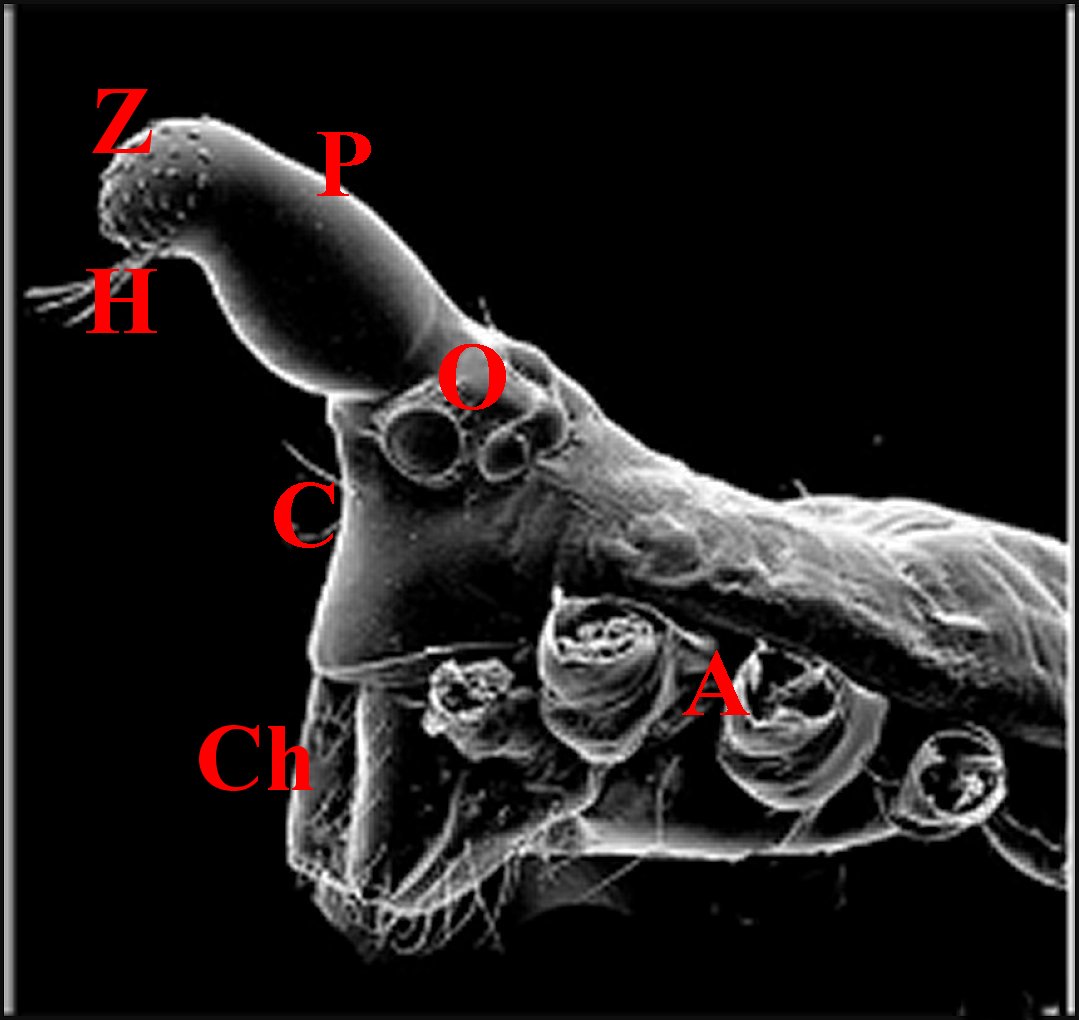
A,
pattes
ambulatoires sectionnées - C, clypeus - Ch,
chélicères -H, poils spécialisés - O, yeux
- P, protubérance ou tubercule interoculaire avec son champ ou
zone de pores (Z)
|
||
Deelemanella est un genre monospécifique représenté par la seule D.borneo Yoshida 2003 découverte sur le mont Kinabalu au Sarawak (Malaisie). Son mâle présente un clypeus saillant mais tronqué, pourvu de longs poils et surtout, une protubérance dorso-médiane effilée et procurvée, siégeant entre les yeux médians (Fig.63). Cette disposition rappelle beaucoup, une nouvelle fois, la "tourelle" d' Argyrodes cognatus (Fig.3),de sorte que l'espèce peut être rattachée au morphotype acuminé.
 |
|
Fig.63 - Deelemanella
borneo, mâle, vue latérale gauche.
D'après
Yoshida
(schéma)
|
| A, pattes ambulatoires sectionnées
-C, clypeus - L, pédipalpe -O, yeux - P, protubérance
procurvée |
Neospintharus est un genre d' Araignées Theridiidae décrit pour la première fois par H. Exline (1950), mis en synonymie avec Argyrodes en 1962, puis enfin revalidé en 2004. La femelle (Fig.64), à abdomen plus ou moins incurvé, tisse une toile de capture irrégulière ,et un cocon sur le modèle des Argyrodes. Le mâle (Fig.65,66) a adopté un morphotype rostré car il est pourvu d'une protubérance oculaire, d'une bosse frontale garnies de poils et d'un sillon ou échancrure séparant ces deux saillies. Des pores excréteurs, indices d'une glande acronale sous-jacente, y sont perceptibles au M.E.B (Fig.66).
 |
 |
 |
| Fig. 64 -
Neospintharus trigonum, femelle, vue latérale gauche avec
l'abdomen dressé puis incurvé. D'après J.H.Emerton (schéma) |
Fig.
65
- Neospintharus
trigonum, mâle, vue latérale
gauche avec ses deux saillies et leurs poils. Schéma. D'après J.H.Emerton (schéma) |
Fig.66- Neospintharus trigonum mâle, vue latérale gauche avec ses deux saillies et leurs poils. D'après Agnarsson(M.E.B.). |
|
A,
pattes ambulatoires sectionnées - Bf, bosse frontale - C,
clypeus - Ch, chélicères -E ou S, échancrure ou
sillon - H,poils - O, yeux - Po, protubérance oculaire
|
||
Faiditus est un cinquième
genre d'Araignées Theridiidae
décrit pour la première fois par Keyserling(1884)
et renfermant, du moins jusqu'en 2017, 59 espèces, toutes
américaines, à l'exception de l' asiatique F. xiphias (Myanmar, Iles
Nicobar, Japon, Krakatau). Mais on peut se demander, d'après des
photos internet de Tanikawa (prises à Okinawa) si ce taxon, par
son mâle, ne se rattache pas au type
nasuté par sa protubérance céphalique (bosse
frontale) en "nez" (Fig.68), proche de celle d'Argyrodes nasutus
(Sri-Lanka)(Fig.10) incluant
vraisemblablement une glande
acronale et, de ce fait, plutôt,
aux Argyrodes
"vrais" dont les femelles adoptent une attitude
de repos semblable
à la sienne (Fig.67).
 |
 |
| Fig.67 - Faiditus xiphias,
femelle au repos. Vue latérale droite. D'après Tanikawa (photo) |
Fig.68- Faiditus xiphias,
mâle se déplaçant. Vue latérale gauche.
D'après
Tanikawa (photo)
|
 |
 |
| Fig.69
- Faiditus
dracus, mâle, vue
latérale gauche D'après Alvarez-Padilla (photo) |
Fig.70
- Faiditus
sp., mâle, Porto-Rico, vue latérale gauche D'après Islandbiogeography.org/ (photo) |
 |
 |
 |
|
Fig.71 -
Faiditus
cancellatus,
mâle, en vue latérale droite.
D'après
Ashley Bradford,
inaturalist.org
|
Fig.72. - Faiditus
chickeringi,
mâle, prosoma, en vue latérale gauche,après section
des pattes ambulatoires.
D'après
Agnarsson (M.E.B.)
|
Fig.73 -Idem, vue
frontale,
D'après Agnarsson (M.E.B.) |
|
C,
clypeus - Ch, chélicères - M,
"menton" avec un champ de
pores - O, yeux
antérieurs et latéraux - P, pattes sectionnées
- S,
sillon garni de poils.
|
||
Au
vu de ce qui précède, il
est à peu prés acquis que la glande clypéale
ou acronale est un caractère princeps d'anatomie interne
caractérisant la sous-famille des Argyrodinae sensu lato
4-3-b- Par
ailleurs,
il est bon de rappeler, comme
déjà
signalé dans l'introduction,
que des glandes propres aux
mâles se rencontrent aussi en dehors des Theridiidae
(Thymoites
par exemple
excepté?....), notamment dans la grande famille des Linyphiidae
où l'on observe de
remarquables modifications de la région clypéale,
notamment chez Bolyphantes
alticeps (Linyphiinae)
et une quantité extraordinaire d' Erigoninae à
protubérances et autres reliefs d'une diversité
inouïe dans la zone "privilégiée" du clypeus ou
bandeau (A.Lopez,
1990b) et la région
oculaire.
Shou-Wang
& al.
(2021) les ont
explorées récemment dans
une remarquable étude tomographique aux rayons X,
sans pour autant la compléter de confirmations
histologiques, une lacune d'autant plus regrettable (Note 1)
que leur "gustatory" ou "nuptial-gift-producing gland", non
présentée en détail, pourrait correspondre, du
moins en partie, à des glandes
gnathocoxales dont le dimorphisme sexuel est prouvé chez
les Erigoninae.
Notes infra-paginales
Note
2: Laboratoire CNRS, Moulis
09200 : fixation des parties étudiées au
glutaraldéhyde à 2,9% dans le tampon Millonig 0,2 M, au
cacodylate pour Kaira
alba),
post-fixation au tétroxyde d’osmium à 0,2% dans le
même tampon, et inclusion en épon (au Spurr pour Kaira
alba) ; coupes fines au microtome Reichert OM U2
contrastées par l’acétate d’uranyle, le citrate de plomb
et examinées ensuite sous 50 KV, au microscope Sopelem du
Laboratoire souterrain. Travail effectué avec
Lysiane Juberthie-Jupeau, directeur de recherches au CNRS,
et ses collaboratrices.
Note 3 : Emerit (2002), qui a
vraisemblablement
capturé son
"Argyrodes
rostratus" en
battant la végétation
xérophile de garrigue établit une confusion facheuse avec
l'espèce du même nom ( Argyrodes
rostratus Blackwall,
1877), taxon tropical
endémique des Seychelles).
LIENS EXTERNES
BIBLIOGRAPHIE
Agnarsson,
I., 2004.- Morphological phylogeny of cobweb spiders and their
relatives (Araneae, Araneoidea, Theridiidae), Zoological Journal of the
Linnean
Society 141 (4), pp. 447-626 : 468-469
Berland,L.,1932.-
Les Araignées in Les
Arachnides, Encycl.entom., XVI, Lechevalier édit., p. 79-404.
Legendre,R., 1960.- Ann.Sci.nat, Zool., 12, p. 507-512.
Lopez,A.,1977b.- Contribution à
l’étude des caractères sexuels somatiques chez les
mâles
d’Aranéides. Thèse doct.Etat es. Sciences,
Univ.Sci.Tech.,
Montpellier, Avril 1977, 117 pp.
Lopez,A.,1987b.-
Newsl.Br.arachnol.Soc.49,p.3-4.
Lopez,A.1974(avec R.Legendre) -
Bull.Soc.Zool.Fr., 99, 3, p.453-460.
Lopez,A.,1975 (avec R.Legendre) -
C.R.Acad.Sci.Paris, 281,D, p.1101-1103.
Lopez,A.,1977.- Bull.soc.zool.France, 102, n° 3, p.261-266.
Lopez,A.,1979 (avec M.Emerit)-
Rev.Arachnol., 2 (4), p.143-153.
Lopez, A.,1980a.(avec C. Juberthie) -
Rev.Arachnol.,3 (1), p.1-11.
Lopez,A.,1980b.(avecM. Emerit,
&L.Juberthie-Jupeau)-Proceed.8. Intern.Arachnol.Kongr.,Wien, 1980,
p.309-313.
Lopez,A.,1981 (avec M. Emerit,) -
Bull.Br.arachnol.Soc.(1981) 5 (4), p.166-168.
Lopez,A., 1990.- Bull.Soc.Sciences nat., 67,
p. 13-22.
Lopez,A.,1990 b - Les caractères
du dimorphisme sexuel chez les mâles d 'Araignées :
revue,données nouvelles et classification . Bull. Soc. Archéol. Scient. Litt.
Béziers, 7e ser., I, p. 43-p. 56.
Millot,J.
1968.- Ordre des Aranéides in Traité de Zoologie,
P.P.Grassé édit., Masson, p. 589-743.
Noirot,
Ch.
& A.Quennedey, 1974.- Ann.Rev.Entomol.,19, p.61-80.
Shou-Wang,L., Lopardo
L. & G.Uhl, 2021 -Diversification
through gustatory courtship: an
X-ray micro-computed tomography study on dwarf spiders.
Frontiers
in Zoology, volume 18,
Article number: 51 (2021),1260
Taylor, H.H. & A.D.Blest, 2009 - The clypeal glands of Mynoglenes and some other Linyphiid Spiders. Journ.of Zoology, Dec.2009, 183 (4), p.473-493.
Whitehouse,M.E.A.,
1987.-
Bull.Br.arachnol.Soc, 7 (5), p. 142-144.
|
RETOUR
|