
Mâle au-dessus de la femelle.
Voir aussi Fig.6 dans
Araign%E9es_et_allomones.html
#Fig.6_-
D'après A.Lopez, 1998.
|
Mastophora
ANATOMIE ET COMPORTEMENT DES ARAIGNEES : VINGT-CINQ ANS DE RECHERCHES Par André LOPEZ, auteur (version 2023) |
| Les Mastophora sont des Araignées Araneidae
propres au Nouveau Monde, d'aspect et comportement
"fantastiques", ne tissant pas de toile orbiculaire mais en revanche
produisant des "globules" visqueux qu'elles manipulent comme les
gauchos leurs "bolas" pour y engluer les proies, exclusivement de
petits Diptères et surtout, des Papillons
Hétérocères mâles. L'attraction de ces
derniers par Mastophora
est sans nul doute liée à un tissu abdominal endocrinoïde
extraordinaire qu'a découvert et nommé
A.Lopez, tissu que les arachnologistes anglophones et hispanophones
semblent avoir ignoré
dans leurs publications concernant un genre
autochtone. ! Un lien avec cet article a néanmoins pu être établi (jusqu'à quand !) depuis https://en.wikipedia.org/wiki/Mastophora_(spider) |
The Mastophora are Araneid Spiders specific to the New
World, of "fantastic" appearance and behavior, not weaving an orbicular
web but on the other hand producing viscous "globules" which they
manipulate like the gauchos their "bolas" to stick the prey, exclusively small Diptera and especially male
Heterocera Moths. The attraction of the latter by Mastophora is
undoubtedly linked to an extraordinary endocrinoid abdominal tissue
discovered and named by A. Lopez, a tissue that the english and
spanish speaking arachnologists seem to
have ignored in their publications concerning a genus that is
nevertheless autochthonous.
|
|
Couleurs
conventionnelles :
En noir et italiques, termes anatomiques ; en violet,,
noms
génériques et spécifiques ; en vert, noms de
familles et sous-familles ; en
orange,, parties
les plus importantes
et résumés ; en bleu,
liens divers
|
|
Abréviations
conventionnelles :
M.E.B. : (photographie en) microscopie électronique à balayage M.E.T. :
(photographie en) microscopie
électronique à transmission
C.H. : coupe histologique
(microscopie photonique)
|
1- Introduction
2- Description
3- Comportement
4- Anatomie
interne
4-1- Glandes à venin
4-2- Glandes gnathocoxales
4-3- Diverticules intestinaux
4-4- Glandes
tégumentaires
4-5- Palpe mâle
4-6- Appareil génital
(abdomen)
4-7- Appareil
séricigène
4-7-a-
Glandes ampullacées
4-7-b-
Glandes piriformes
4-7-c- Glandes aciniformes
4-7-d- Glandes agrégées
4-7-e- Glandes flagelliformes
4-7-f-Glandes tubuliformes
4-8- Tissu folliculaire
endocrinoïde abdominal
4-8-a - Histologie
4-8-b - Ultrastructure
5 - Commentaires
5-1-Considérations
anatomo-physiologiques
5 -1-a
- Appareil génital
5-1-b - Appareil séricigène
5-1-c
- Tube digestif
5-1-d - Tissu
folliculaire endocrinoïde abdominal
5-1-d-a
- Rapports avec la glande de mue
5-1-e-b - Hypothèses
fonctionnelles
5-2 -
Considérations phylogéniques
Créé
par Holmberg
(1876) le
genre Mastophora est
un groupement
d' Araignées Aranéomorphes de la famille des Araneidae.
Appelé aussi Glyptocranium et
Agathostichus
par Simon
(1895,1897), il est placé aujourd’hui
dans la sous-famille des Cyrtarachninae,
y formant la tribu
des Mastophorini, avec
les
genres Cladomelea,
Acantharachne
(Afrique)
et Ordgarius
(Australie,Océanie).
Au
point vue biochimique
et éthologique, il fait partie du groupe étrange des Araignées à
allomones
En ce qui concerne la distribution, toutes
les espèces du
genre Mastophora se
rencontrent exclusivement en
Amérique tempérée et surtout tropicale.
Au point de vue étymologique et par
allusion à la forme des excroissances
somatiques, le nom "Mastophora" pourrait avoir
été construit à partir du grec ancien
: ''mastós''
(« mamelle, sein
») et φορός,
phoros
(« porteur »)
L'anatomie
et le
comportement des Mastophorini offrent
des
particularités uniques dans
l'ordre des Araignées, peut être même
dans le monde
animal tout entier.
Comme
les autres Mastophorini, les Mastophora se
singularisent par un corps trapu et
des pattes épineuses.
Le
céphalothorax
montre de curieuses protubérances
dorsales, soit une paire de
«cornes
occipitales» plus ou moins bifurquées (Fig.1a,5,6), soit
des files de pointes
acérées. Leur abdomen
est globuleux, avec des bosses
dorso-latérales parfois très saillantes et des
teintes cryptiques. Le dimorphisme
sexuel est
considérable (mâles
"pygmées") (Fig.1b)
|
|
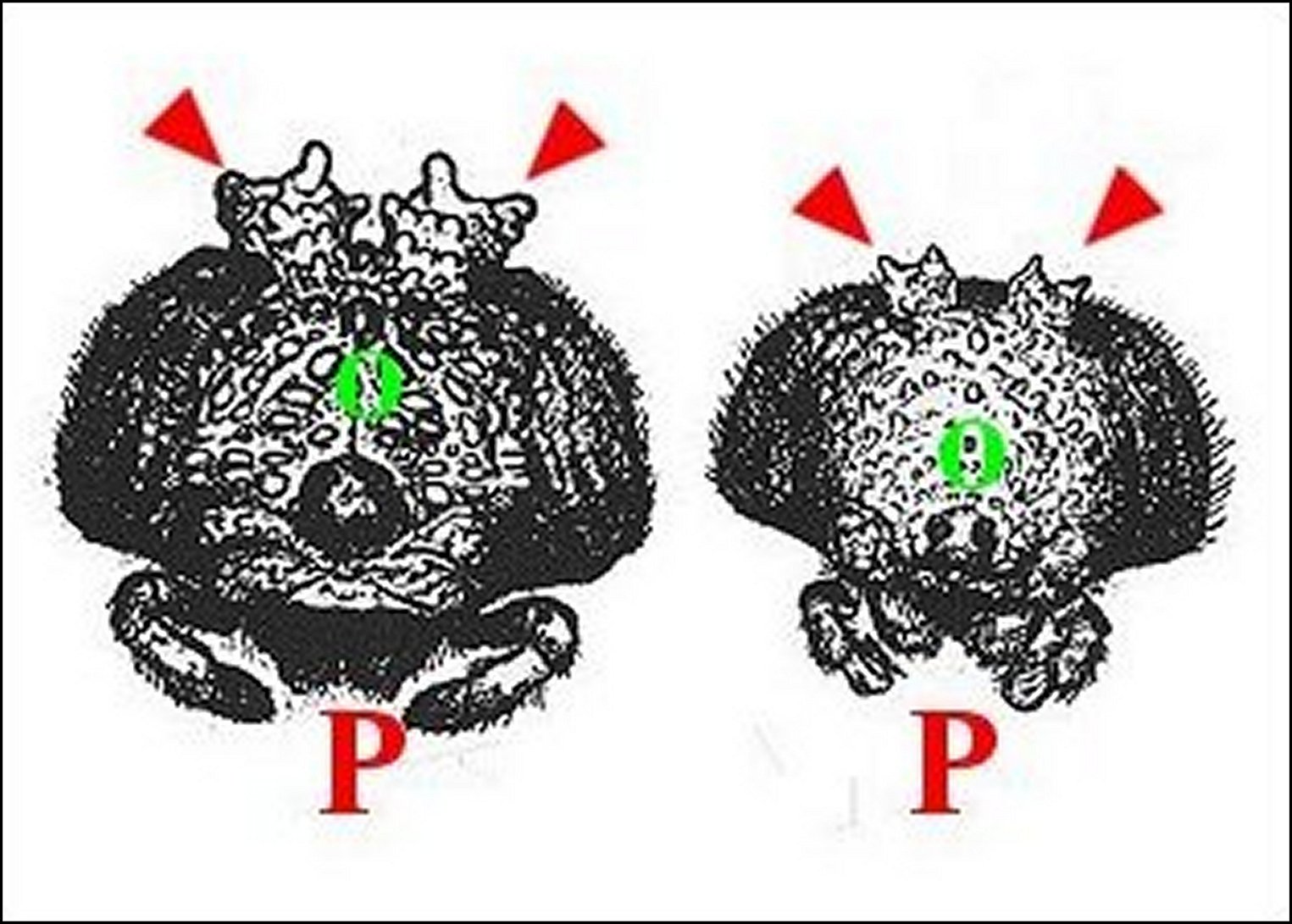
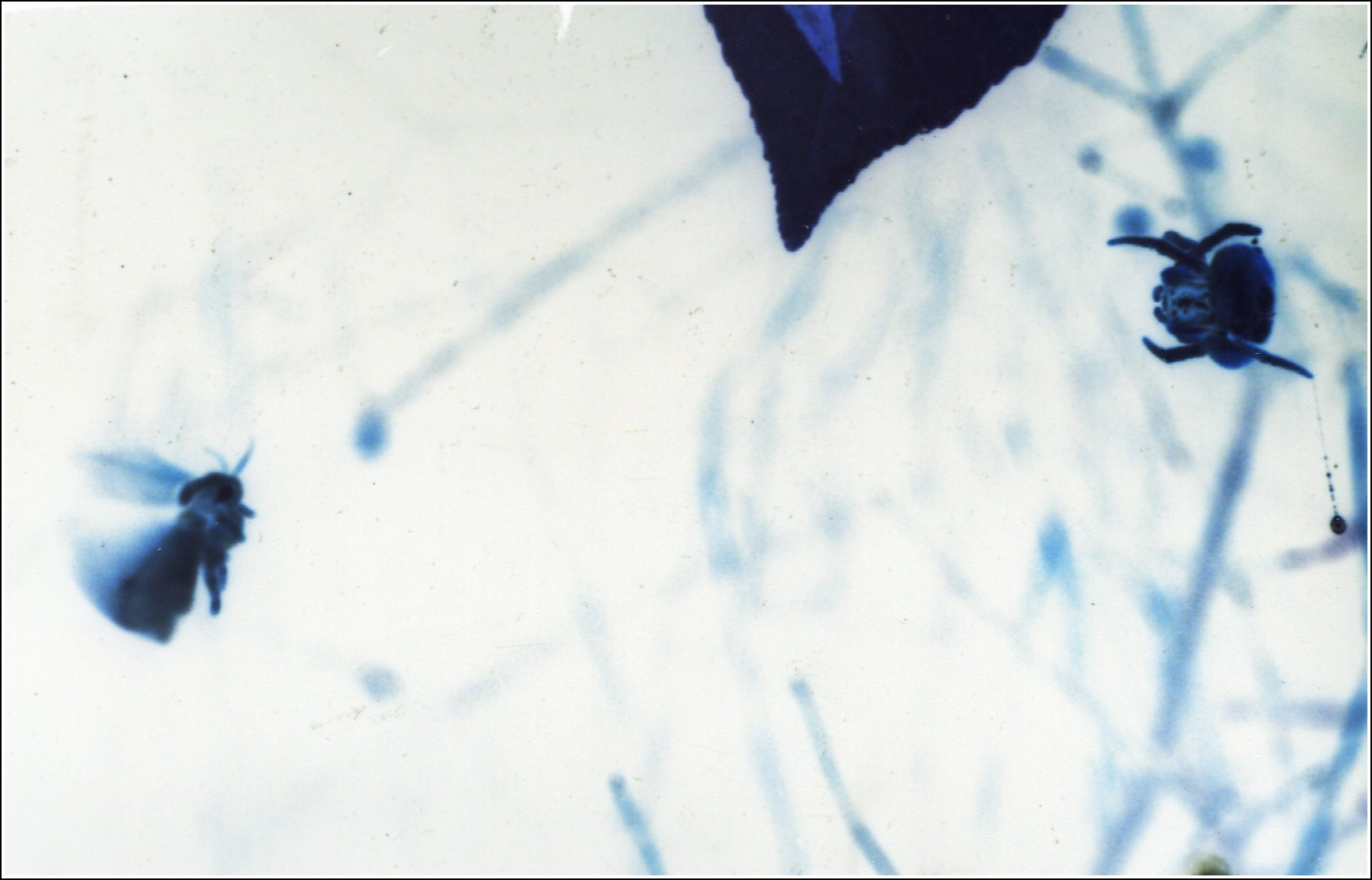
| Fig. 1 a - Mastophora cornigera, à gauche et M.bisaccata, à droite : céphalothorax de femelles vus de face. | Fig.
1 b - Mastophora bisaccata, couple. Mâle au-dessus de la femelle. Voir aussi Fig.6 dans Araign%E9es_et_allomones.html #Fig.6_- |
| F,
femelle - M, mâle nain - O,
yeux -
P, pédipalpes.
Flèches : "cornes". D'après A.Lopez, 1998. |
Les Mastophora immatures
et les mâles subadultes capturent
uniquement des Diptères Nématocères Psychodidae (Yeargan
& Quate, 1996,1997)
et
les femelles, des Papillons
Hétérocères (Gertsch,1947;
Stowe,1986).
Tous ces
Insectes sont des mâles que
l'araignée attire en
émettant des
substances volatiles odorantes qui
simulent les phéromones sexuelles
de leurs femelles et font donc partie des allomones. Pour
cela, les femelles ne tissent pas de toile
orbiculaire mais un simple fil horizontal de suspension (Fig.3, 4) et
un "bola"
pendant (Fig.2,3), ce dernier ainsi
nommé car évoquant le
dispositif de capture
du bétail utilisé par les gauchos sud-américains pour entraver ses pattes.
Il est homologue de la spirale
adhésive des
Araignées orbitèles et
correspond donc bien à un "fil de capture gluant". Il se compose
d'un
fil vertical et d'un globule visqueux dont l' adhésivité,
le caractère
extensible et la richesse en eau sont extrêmes (Eberhard,1980) mais
qui se dessèchent rapidement pour perdre leurs
propriétés, ce qui expliquerait
que l' Araignée les ingère et les reconfectionne toutes
les 15 à 30 minutes.
Mastophora
manipule
le bola avec sa deuxième paire
de pattes (PII) en lui
imprimant un
mouvement de
rotation (Fig.3) et englue
à la volée les proies qu'elle
a attirées, le globule
adhérant au tégument des Lépidoptères malgré
la couverture d'écailles protectrices (Fig.4). Elle les hale
ensuite avec le
fil ou descend vers elles, les
saisit entre les pattes, les paralyse
avec
ses chélicères
et
les enveloppe enfin de soie avant de les consommer.
Les jeunes Mastophoras des deux sexes sorties du cocon et les mâles émettant une "odeur" attractive pour les Diptères Nématocères, saisiraient directement ces derniers avec leurs deux premières paires de pattes, PI et PII, garnies de "herses" d' "épines" (Fig.5) sans utiliser de bolas (Eberhard,1980) ou en confectionnant des "miniatures" (Stowe,1986) , ce qui reste à démontrer.
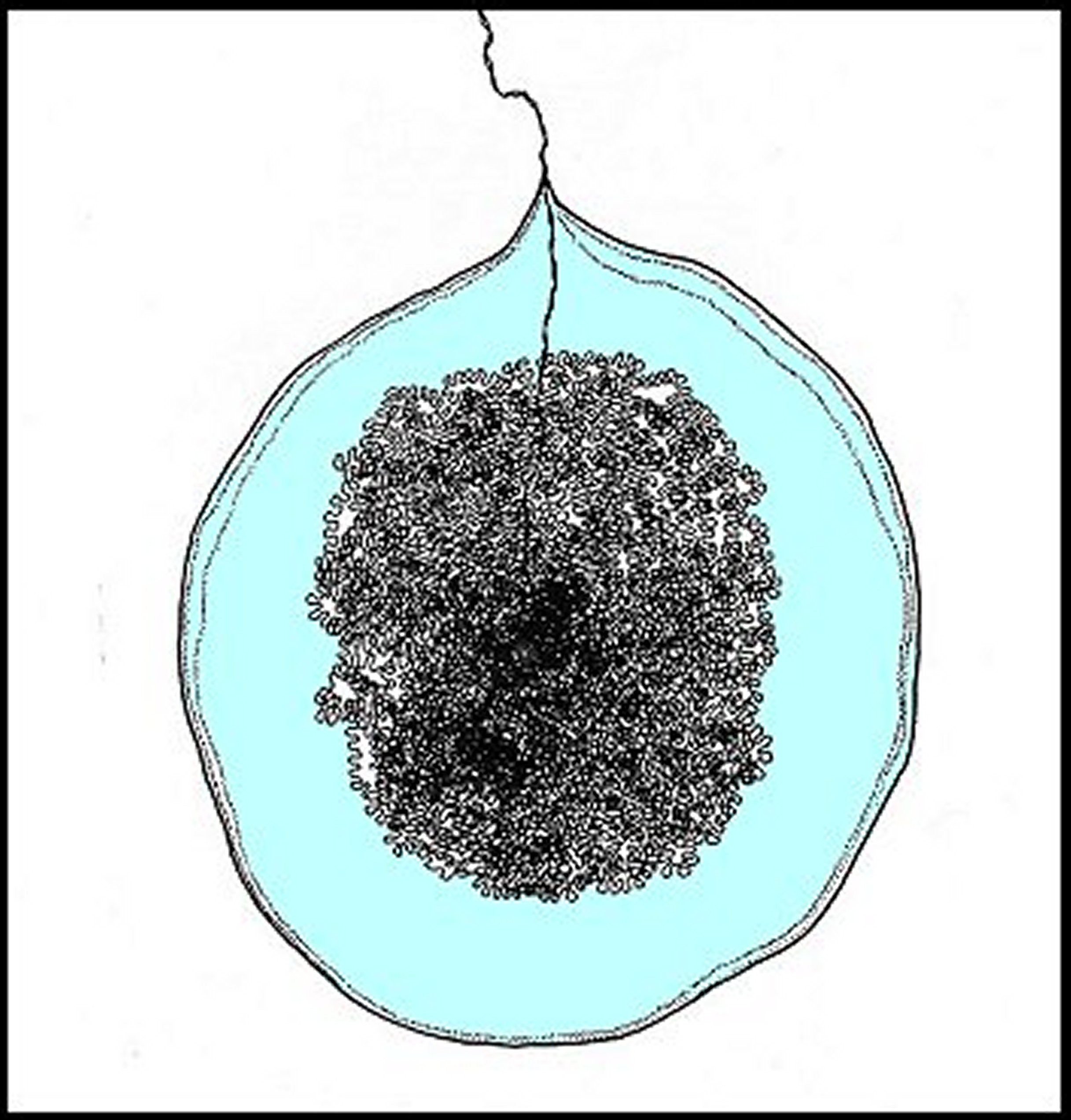 |
| Fig. 2 - Bola montrant le fil de suspension, pelotonné en son centre, et la glue qui l'entoure. |
 |
 |
| Fig.3a
- Mastophora bisaccata femelle
tenant son bola avec
une patte (à droite) et attirant un
Hétérocère mâle (à gauche). Floride. D'après une diapositive. |
Fig.3b - Les mêmes. Image négative |
 |
|
Fig.4
- Mastophora bisaccata,
même femelle, venant de
capturer
l'Hétérocère mâle qui s'est englué
sur le bola.
|
|
D'après
une diapositive.
|
 |
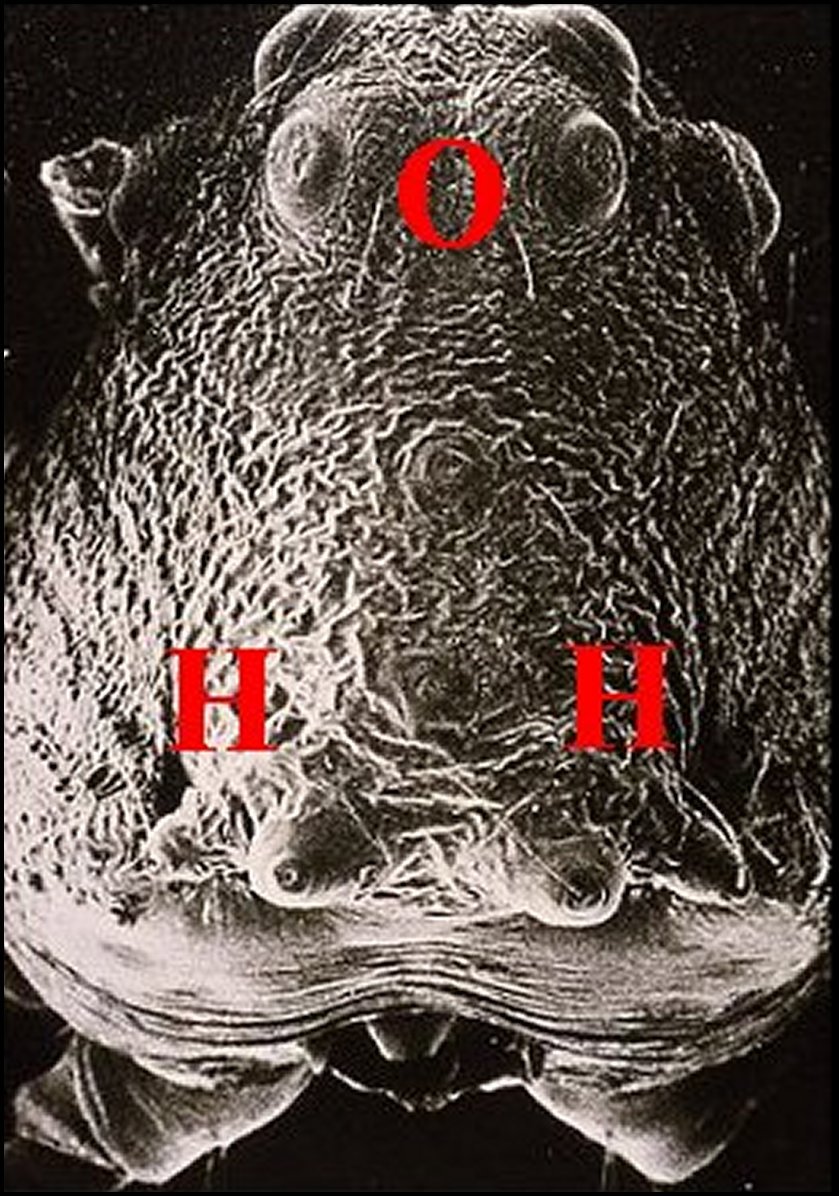 |
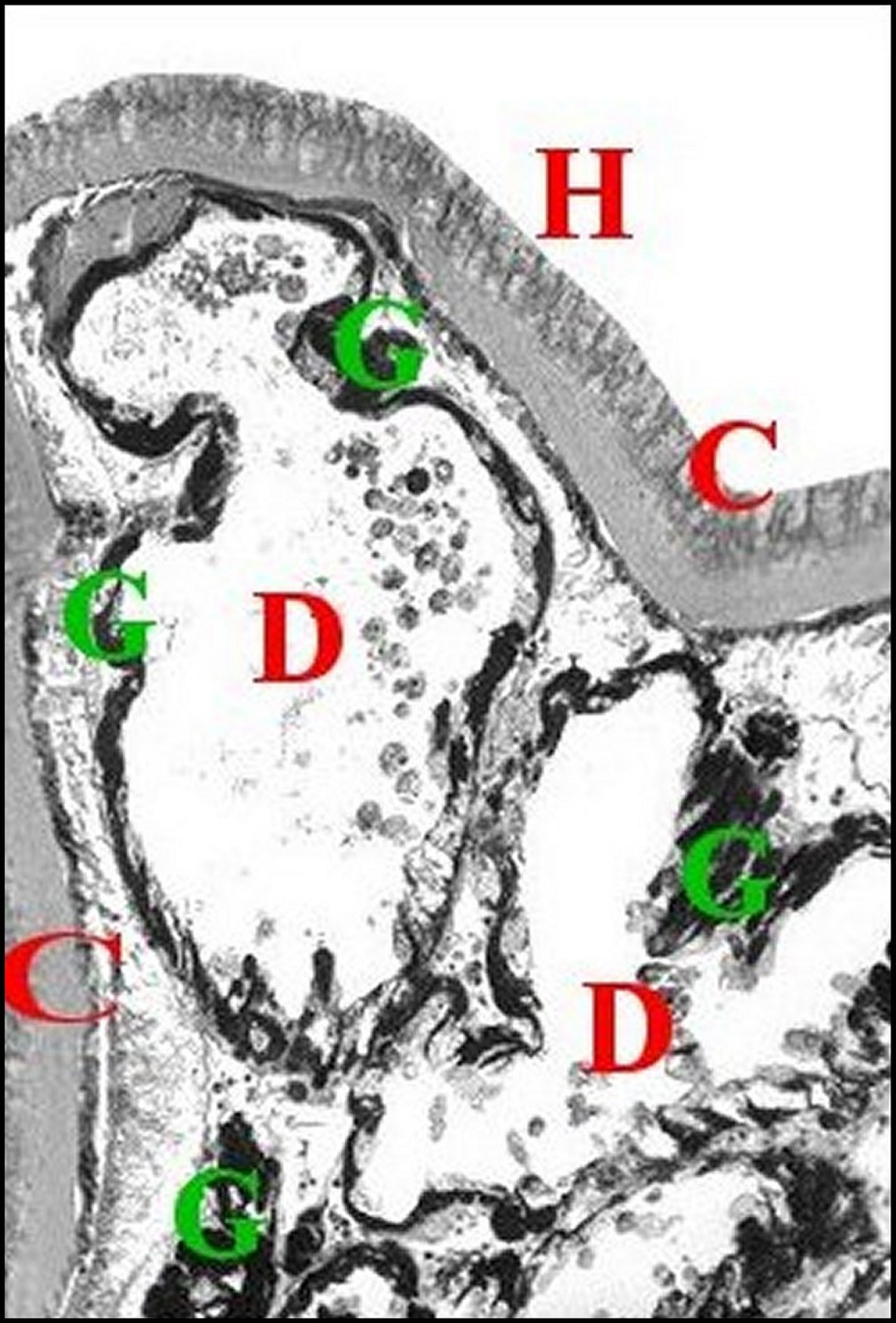
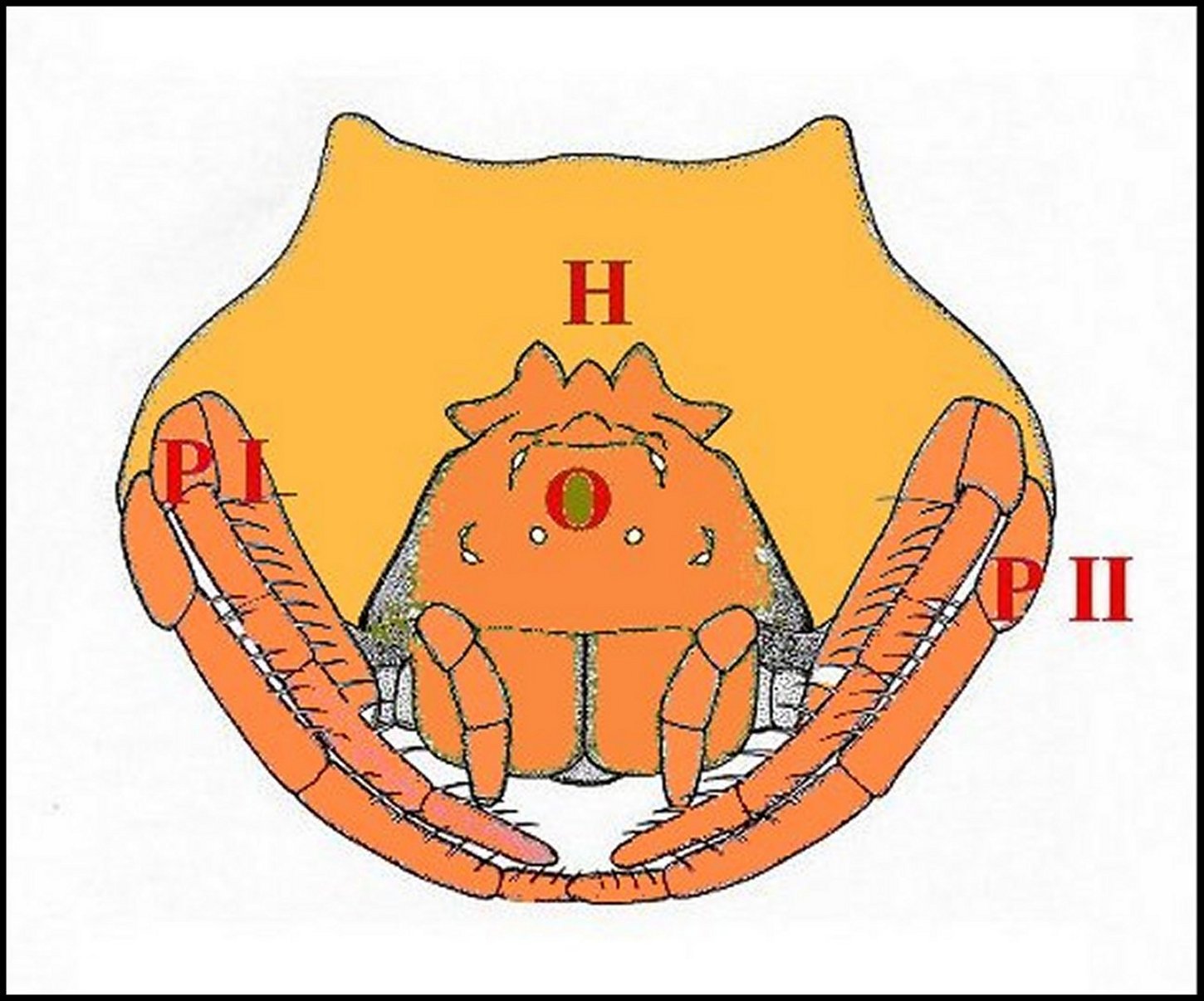
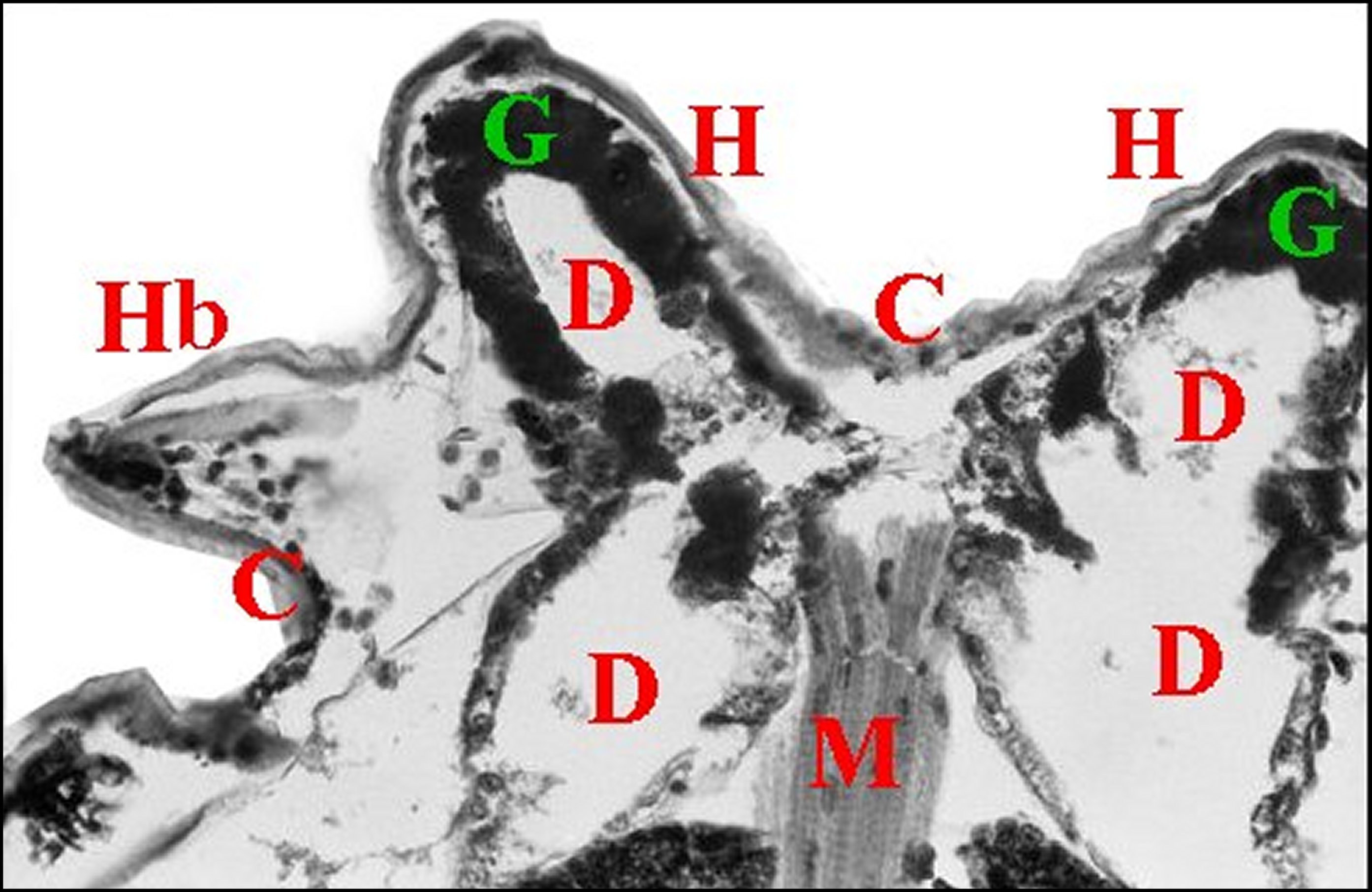
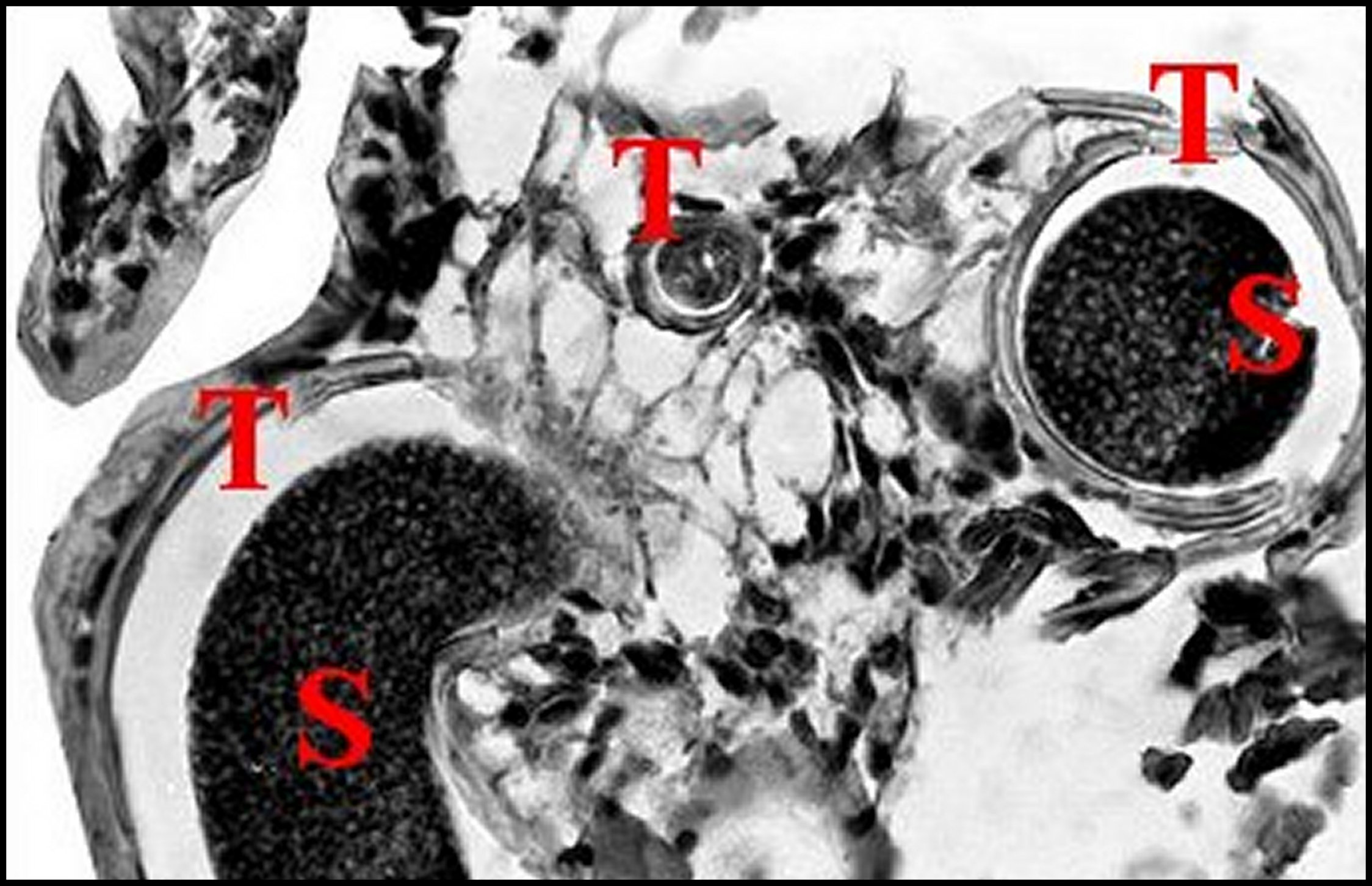
| Fig.5 - Mastophora immature, vue de face : céphalothorax avec ses cornes, pattes I et II, abdomen avec ses bosses à l'arrière plan . | Fig.6 - Mastophora cornigera , mâle : vue dorsale de la carapace montrant les deux protubérances postérieures ou "cornes" (H). |
| H, "cornes" - O, région oculaire- PI et PII, pattes hérissées de soies. Dessin. | 0, région oculaire (©A.Lopez, M.E.B.) |
Comme pour celle de Kaira
(1986,1998, 2001),
rivale de Mastophora
en
"extravagance" structurale et éthologique, Lopez (1985a,1985b) a effectué sa toute
première étude jamais effectuée
au microscope en
Arachnologie. Ses
recherches, de
nature histologique,
ont porté sur Mastophora
cornigera, M.
bisaccata (Mastophoreae)
et, à titre comparatif, sur Celaenia excavata (Celaenieae)
de Nouvelle-Zélande
ainsi que Poecilopachys australasia,
d'
Australie, autres Cyrtarachninae.
Il est
à préciser ici que les
deux premiers
taxons, provenant des
États-Unis (Californie, Floride) où
l’ Américain Stowe
les avait récoltés sans mesurer l’importance
réelle de ce matériel , étaient
représentés par des exemplaires à divers stades
évolutifs, y
compris des jeunes
femelles et tous les
mâles mûrs sortis depuis peu du cocon (M.bisaccata).
Les études microscopiques
de l'auteur français
ont
révélé,
comme chez Kaira, d’étonnantes
structures internes en rapport avec le comportement hors normes des
taxons
concernés, cela au grand dam des arachnologistes
Américains, Stowe inclus, qui
ont
préféré les occulter totalement dans
leur littérature. Selon toute vraisemblance, ces
« scientifiques » US
ont
du même
faire pression sur
« Wikipédia » puis sur
« Free
Wiki »,
où les deux articles correspondants ont
été
stérilisés sans vergogne, tant ils faisaient
ombrage à leurs versions anglo-américaines. En
ce qui
concerne le présent
article sur Mastophora
récupéré avant destruction totale, il
suffit de le comparer avec "Wiki" en utilisant, comme
déjà évoqué sous le titre, les deux liens
suivants,
l'un vers le texte françisé de cette "Encyclopédie" et l'autre vers
l'anglais :
https://en.wikipedia.org/wiki/Mastophora_(spider)l
Elles
montrent un corps allongé et
sacciforme reposant sur le syncérébron
(Fig.11). Il est
formé par un épithélium
sécréteur simple et par une tunique musculaire
contractile à fibres
striées (Fig.11). Le canal
excréteur est dépourvu de toute
différenciation
particulière.
Ces glandes "maxillaires" ou
« salivaires », logées dans les
gnathocoxae, ne
présentent qu’un seul
type de corps
sécréteurs tubulo-acinoïdes, y compris
chez le mâle (Fig.7). Il n'existe donc pas de dimorphisme
sexuel des glandes gnathocoxales contrairement à ce que l'on observe
chez d'autres Araneidae, Kaira alba
comprise.
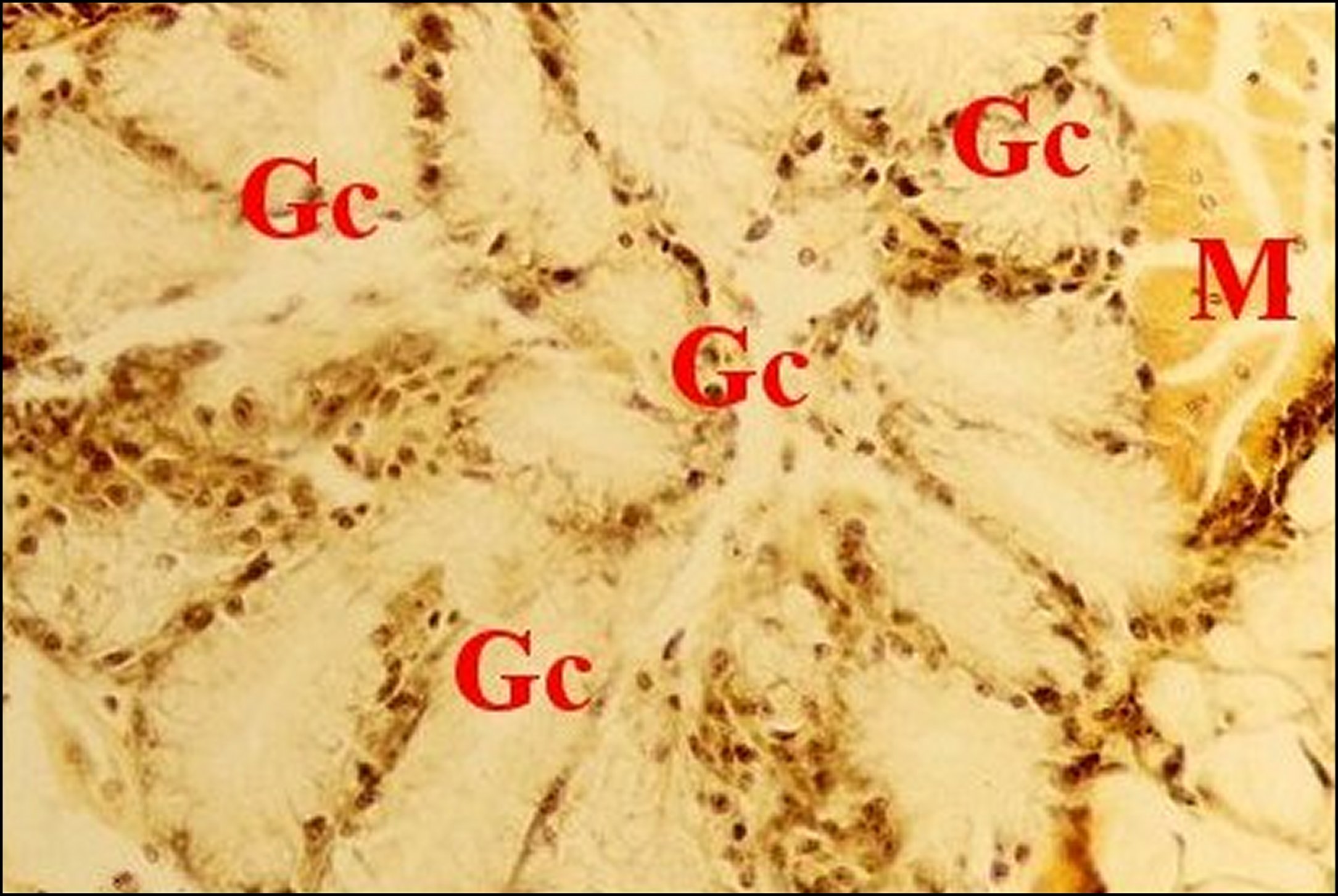 |
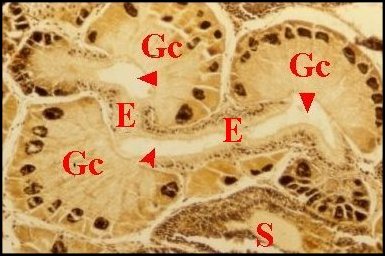
| Fig.7 - Mastophora mâle : coupe histologique de gnathocoxa. |
|
Les
glandes "salivaires" sont toutes "classiques" (Gc). M,
muscles .Comparer avec la Fig.39, Kaira mâle
(©A.Lopez,C.H.).
|
L’intestin
moyen de Mastophora émet
des diverticules
thoracentériques
développés :
expansions latérales,
s’engageant dans les coxae des pattes ambulatoires et surtout,
diverticules du
plan supérieur. Ces
derniers offrent la particularité
singulière de
pénétrer
dans les «cornes» dorsales (Fig.1,5,6), en
occupent la majeure partie (8 à12),
surmontent le cerveau ou syncérébron
(Fig.10,11) et
entrent en contact avec
des muscles striés prosomatiques,
aussi bien chez les femelles
immatures (Fig.10 à
12)
que chez les adultes (Fig.32, Fig.33) et les mâles (Fig. 8,9).
Comme
les diverticules
intra-abdominaux du
chylentéron (intestin), ils comportent à leur surface des
cellules particulières,
décrites pour la première fois par Millot (1931) chez
d’autres Araignées : les
gua(ni)nocytes ou cellules à guanine,
ainsi nommées car
elles contiennent cette
substance, un produit
d’excrétion
cristallin
biréfringent, s’éclairant donc en lumière
polarisée (Fig.9, Fig.33) et qui apparaît
très blanc dans les images négatives. Un sinus
sanguin sépare les
diverticules du tégument dont l’épithélium aplati,
sans différenciation
glandulaire, est recouvert d’une épaisse cuticule verruqueuse
ininterrompue.
 |
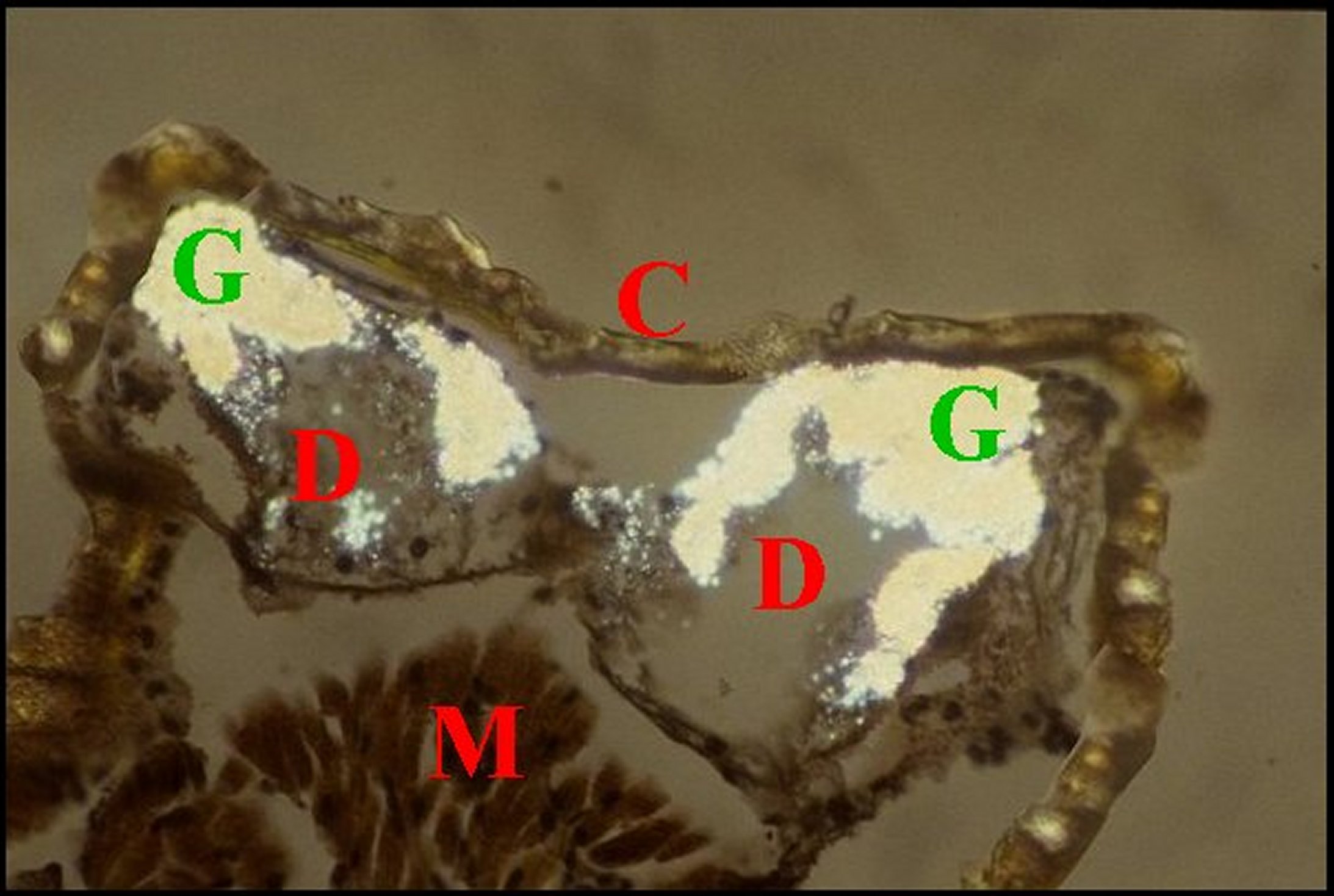 |
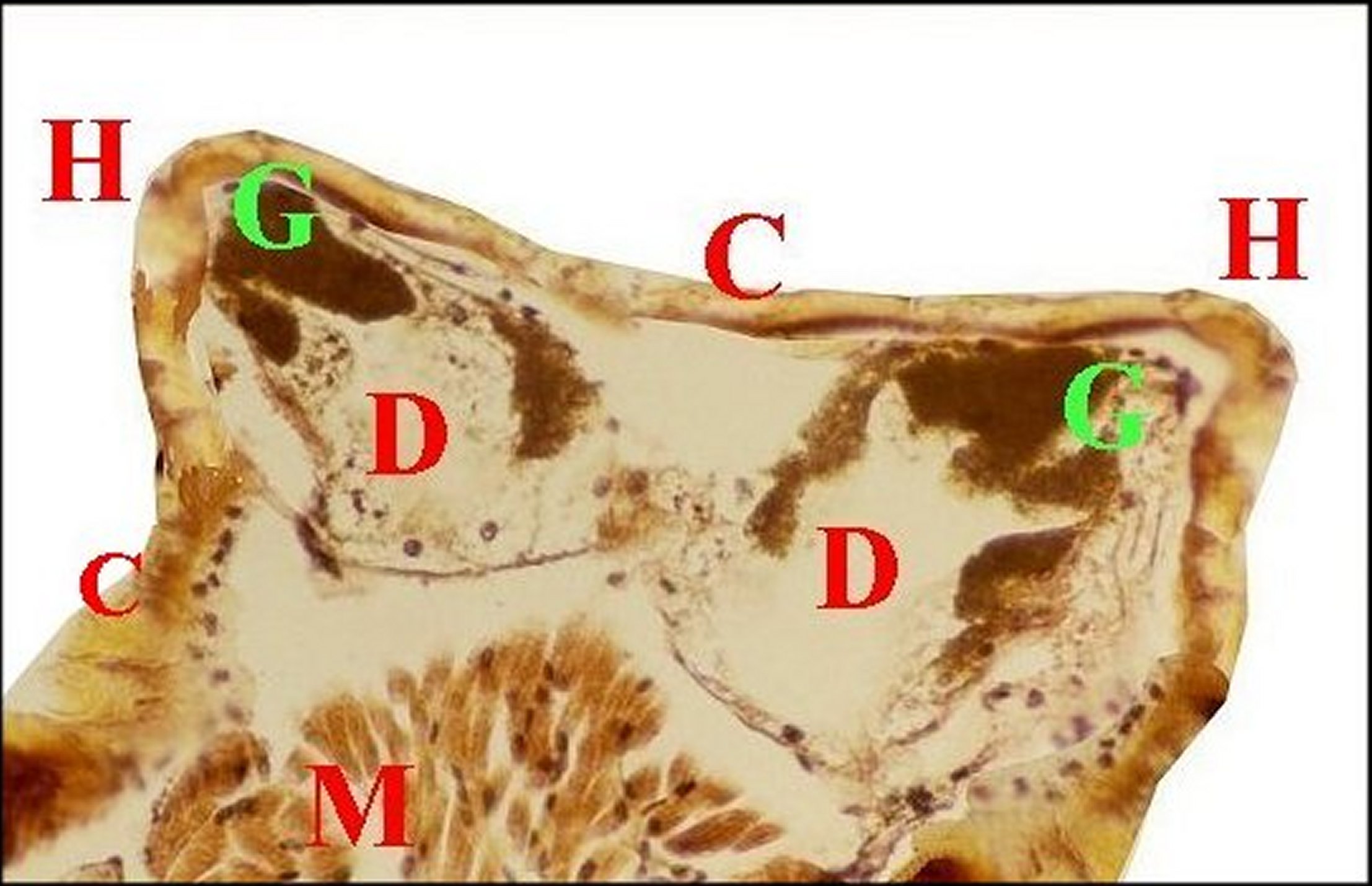
| Fig.8 - Mastophora mâle : cornes céphalothoraciques renfermant des diverticules intestinaux. | Fig.9 -Mastophora mâle, même coupe vue en lumière polarisée. |
| C, cuticule - D, diverticule chylentérique - G, guaninocytes - H, corne - M, muscle (©A.Lopez, C.H.en lumière normale). | C, cuticule - D, diverticule chylentérique G, guaninocytes intestinaux biréfringents - H, corne -M, muscle (©A.Lopez,, C.H.polarisation). |
 |
 |
 |
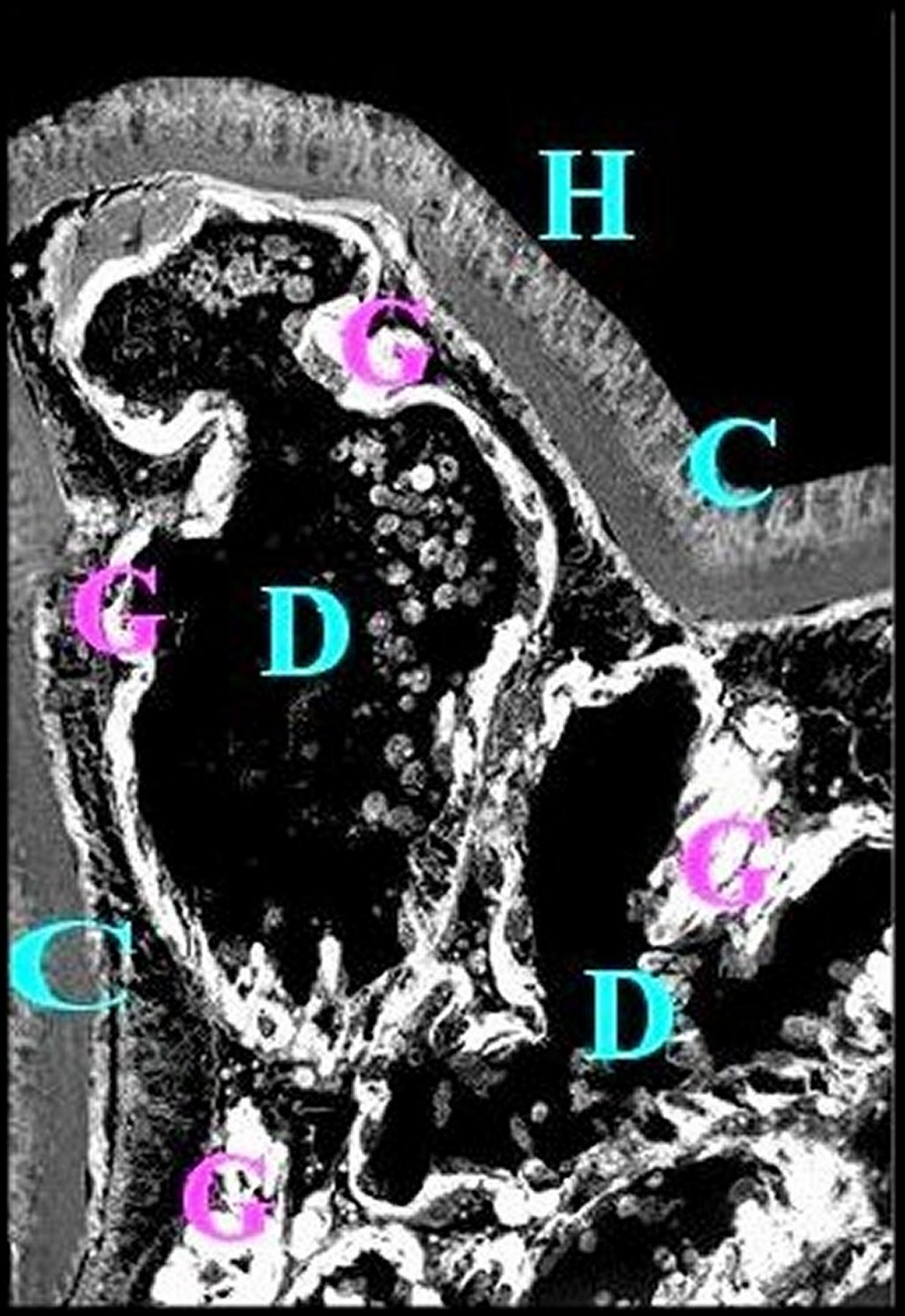
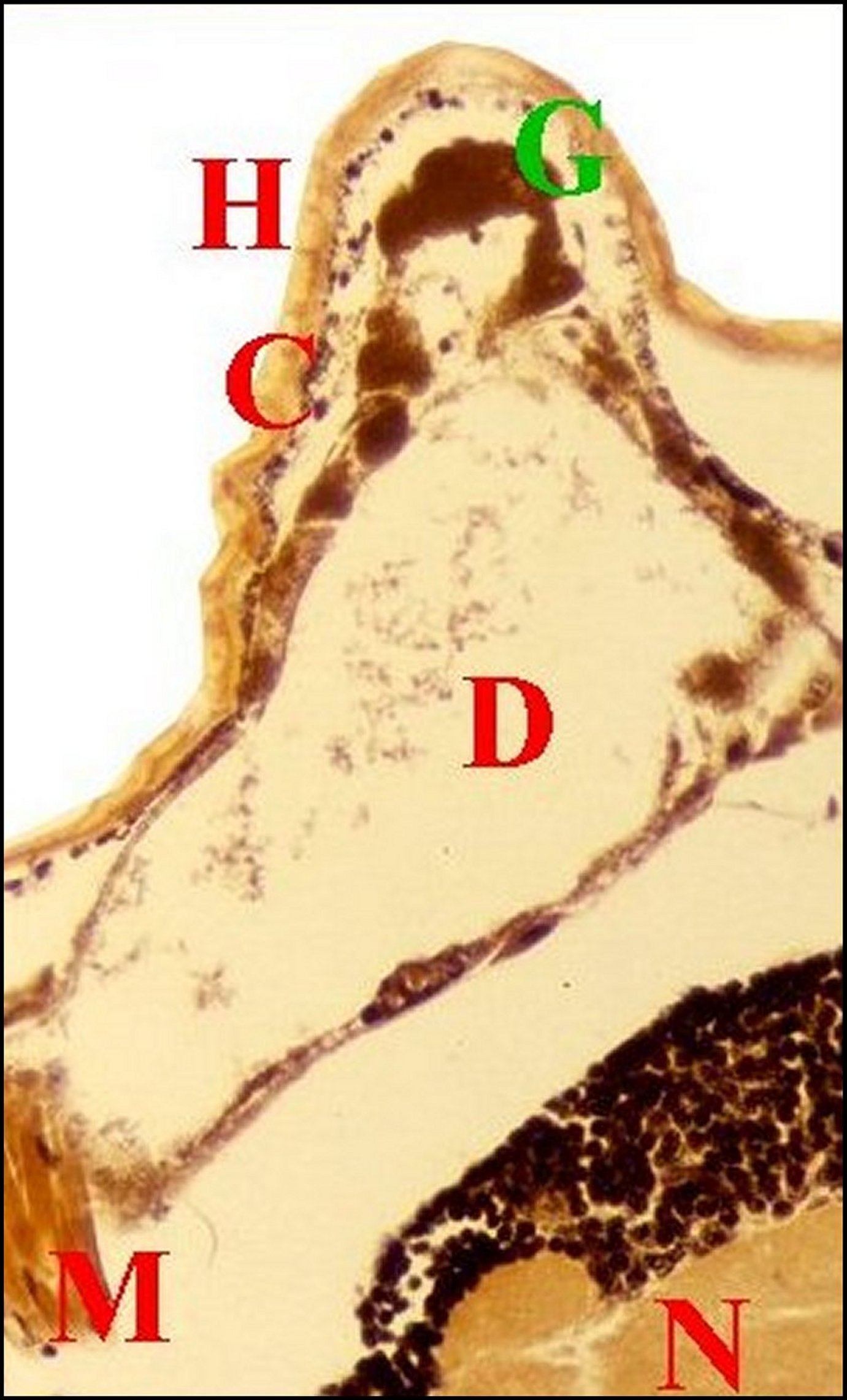
| Fig.10 - Mastophora cornigera, femelle immature : coupe parasagittale du prosoma passant par une corne. | Fig.11- Mastophora cornigera , autre femelle immature : coupe parasagittale du prosoma passant par une corne. | |
| C,cuticule
- Ch,
chélicère -D,
diverticule
chylentérique - G, guanocytes - Gn,
gnathocoxa
- H, corne -
Hb,
autre corne
(bifurquée) -
M, muscle -
N, cerveau
-V,
glande à venin (©A.Lopez, C.H., lumière normale). |
||
Chez
les mâles et femelles immatures
des Mastophora, les pattes
antérieures, garnies d’une
«herse» de
soies prolatérales
(fig.5), renferment de nombreuses glandes
tégumentaires dans leurs
articles. Bien visibles en
coupes histologiques et moins nombreuses
chez le
mâle (Fig13,14) que chez la femelle (Fig.15), elles sont
sous-cuticulaires,
intra-épidermiques et comportent un adénocyte avec
"réservoir", un
canalicule excréteur, sa cellule satellite ou cellule
canaliculaire.
L'adénocyte
est pourvu d’un gros noyau
nucléolé (Fig.14) et d’un "réservoir" ovoïde
(Fig.13 à 15). Ce
dernier, qui
correspond indiscutablement à une cavité extracellulaire
sur le plan
ultrastructural, montre de fines stries acidophiles radiaires, en fait
des microvillosités et
est centré par un canalicule récepteur (fig.14 :
flèche), leur
ensemble formant le classique "appareil terminal" ("end
apparatus") décrit chez les Insectes (Noirot
& Quennedey, 2003)
et est prolongé par un canalicule excréteur. Ce dernier s’entoure d'une ou plusieurs cellules
satellites à noyau
très chromatique et traverse ensuite la cuticule
pour aboutir en
surface à un
pore excréteur
(Fig.13 : flèches).
 |
 |
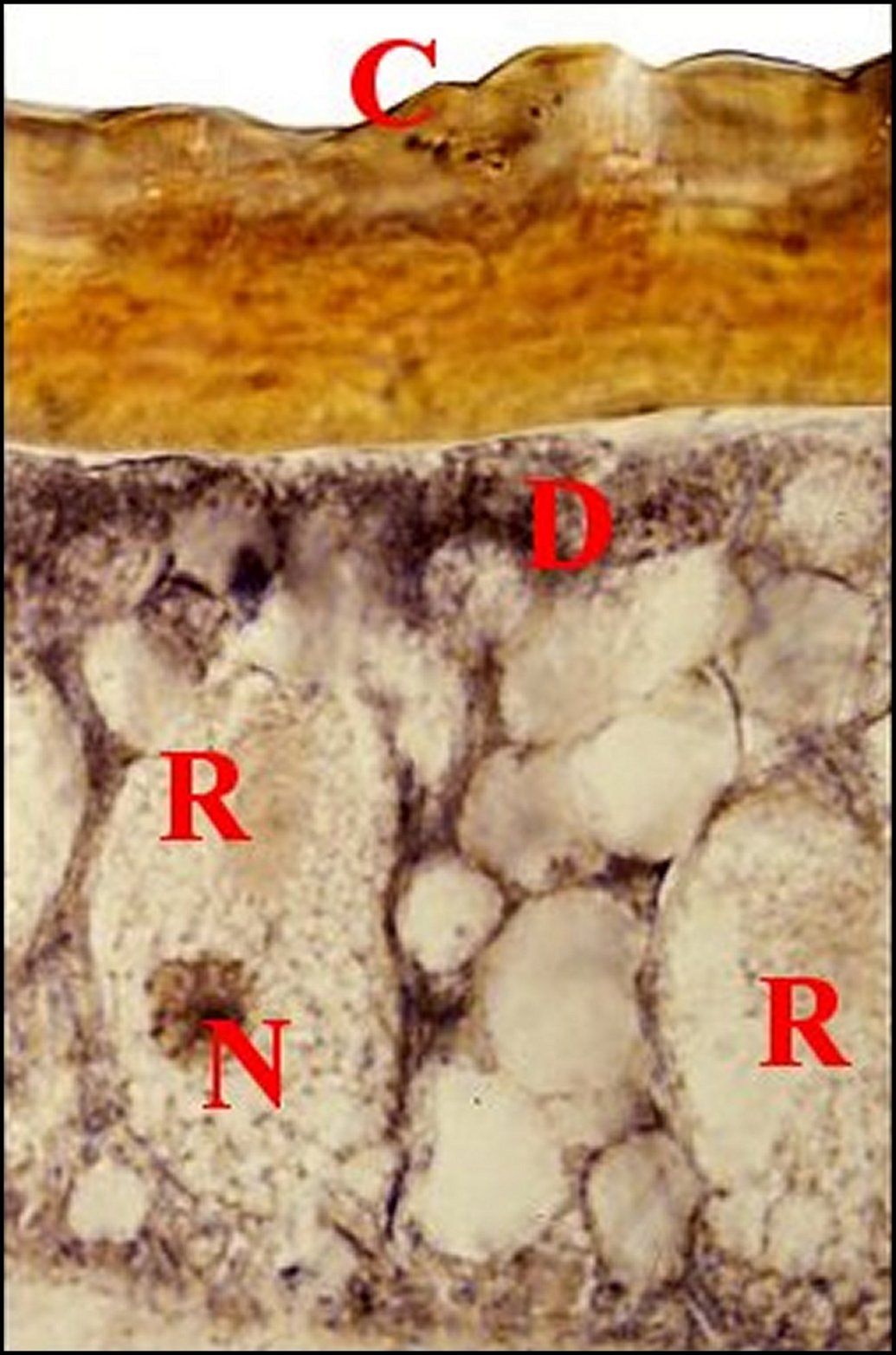 |
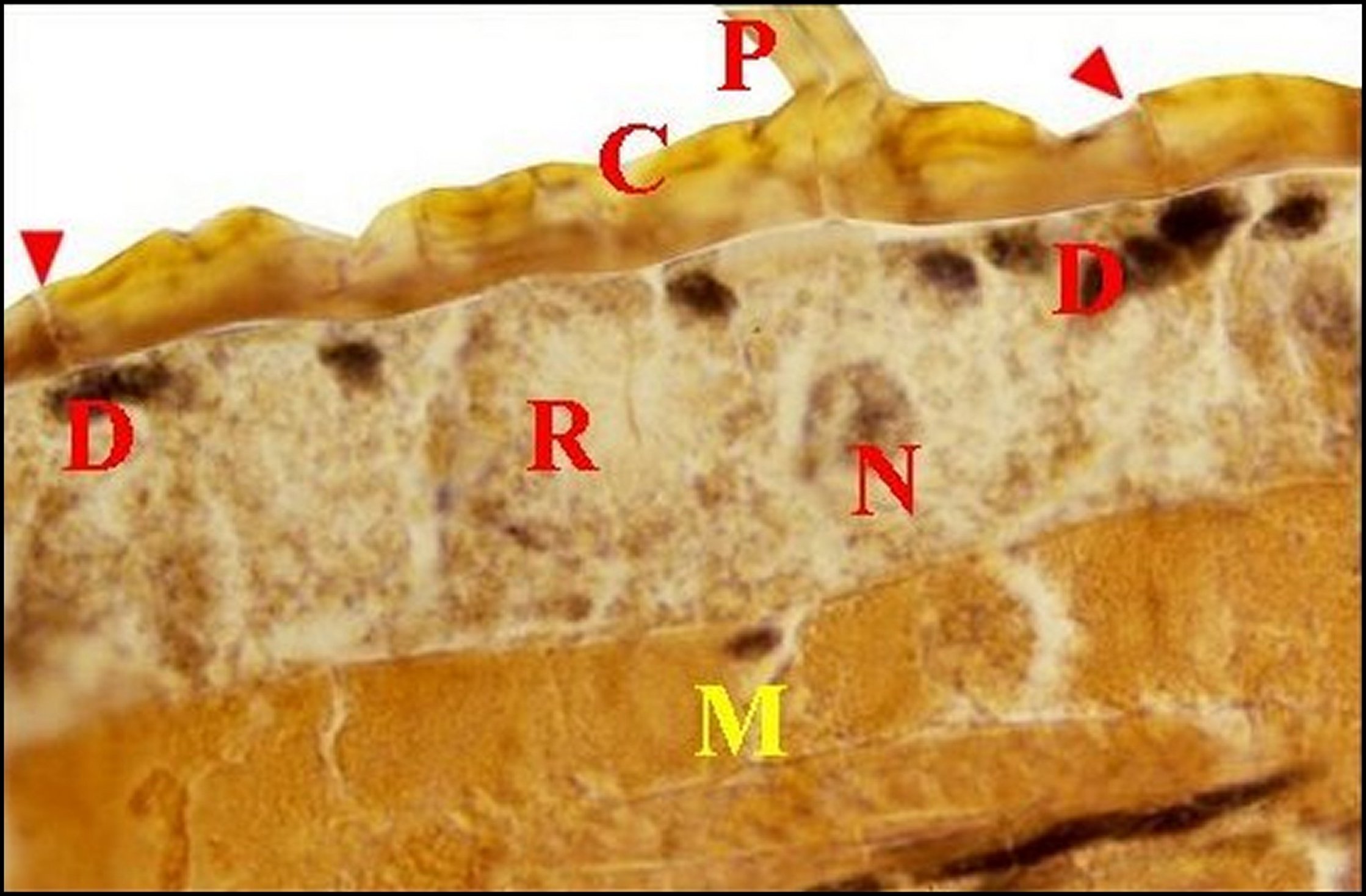
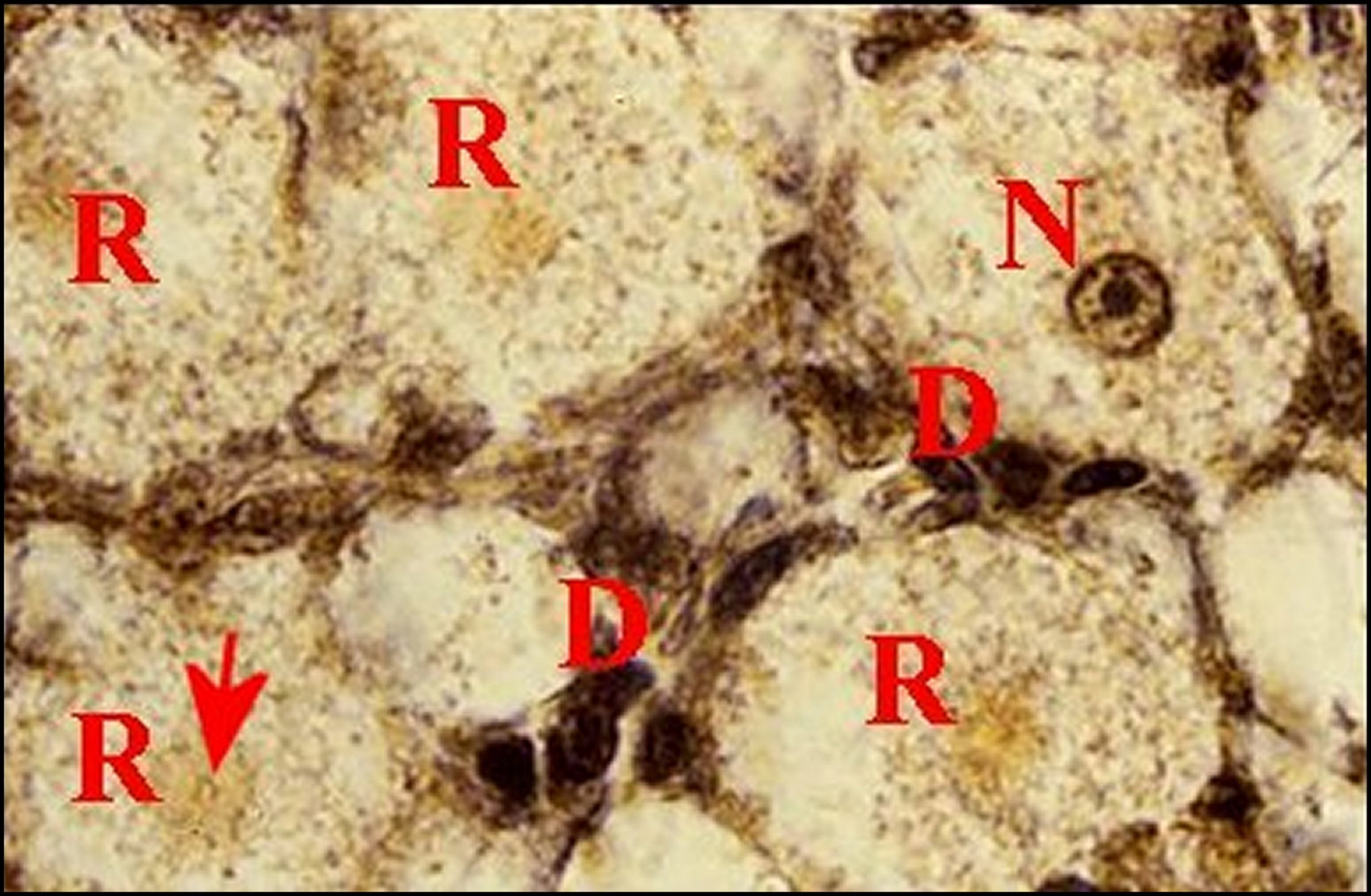
| Fig.13 - Mastophora cornigera, mâle : tégument de patte antérieure en coupe transversale. | Fig.15 - Mastophora cornigera, femelle immature : tégument de patte antérieure en coupe perpendiculaire | |
|
C,
cuticule - D, noyaux des
cellules
canaliculaires - M, muscle - N, noyau d' adénocyte - P, poil -
R,
"réservoir". Flèches : pores de canalicules
excréteurs
(©A.Lopez,
C.H.)
|
||
Chez
le mâle mûr
de Mastophora, issu
directement du cocon - fait
extraordinaire
chez les Araignées où
il ne se retrouverait guère
que dans le genre Arachnura
-
le bulbe du pédipalpe (Fig.16)
englobe un long tube
séminifère.
Son
réservoir montre
une partie chitineuse
fenestrée et un épithélium
glandulaire banal, à une seule
catégorie de cellules
comme chez les autres Araneidae
et
d’ailleurs, comme dans
l’ordre tout entier (Lopez, 1977). Ce réservoir
était rempli de
spermatozoïdes
encapsulés chez tous les mâles
examinés (Fig.16) ;
provenant du tractus génital
après induction spermatique,
ils peuvent y
séjourner très longtemps.
 |
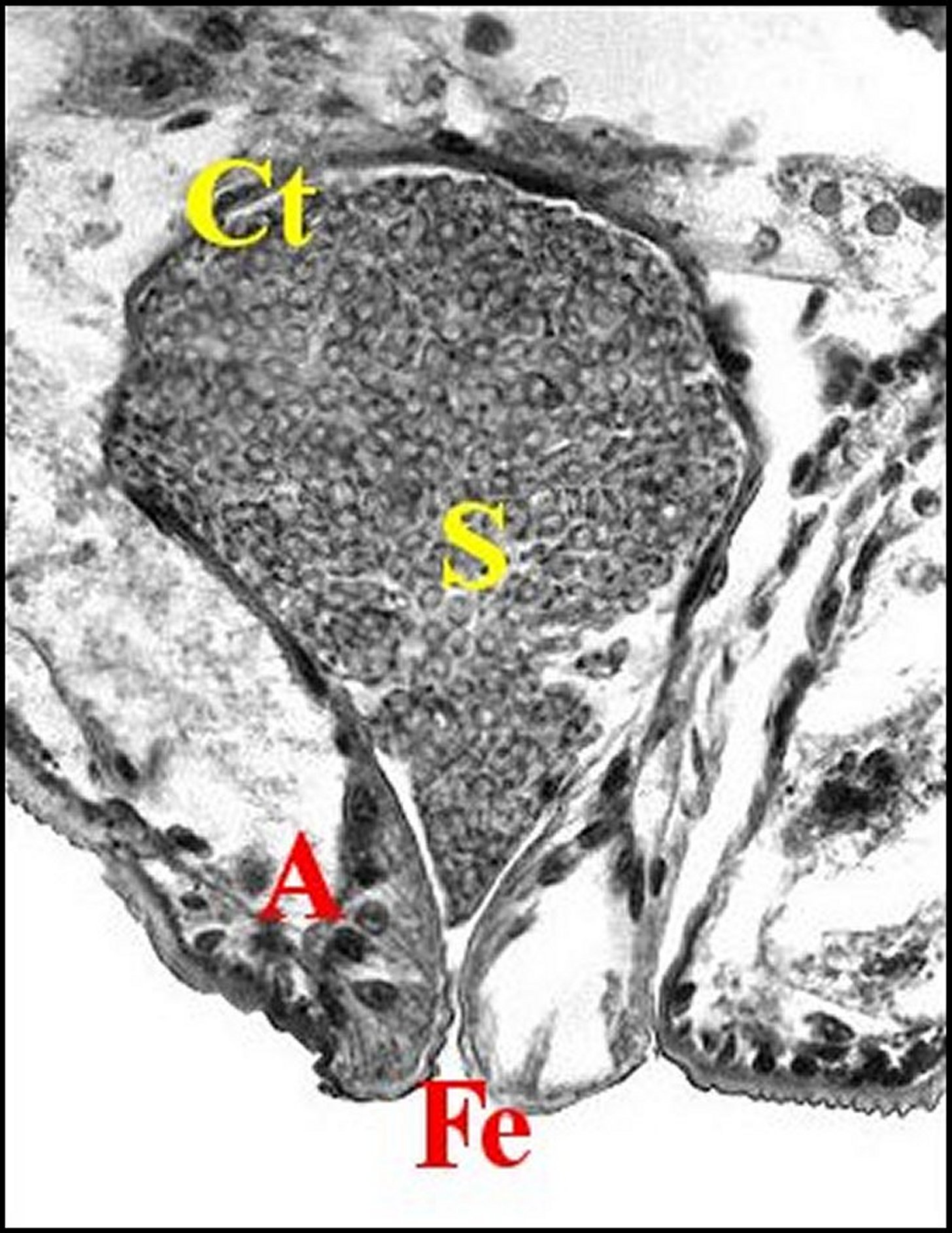
| Fig.16 - Mastophora bisaccata mâle: bulbe après induction spermatique, vue trés partielle. |
| S,
spermatozoïdes
(gamètes) - T, paroi chitineuse du tube séminifère
dont sont visibles trois
sections que sépare la glande du palpe.
(©A.Lopez,
C.H.)
|
4-6
- Appareil génital (Abdomen)
L’abdomen
de Mastophora renferme,
chez la femelle, un appareil
génital sans particularité
notable. Celui du mâle,
en revanche, se singularise, tout comme le pédipalpe, par un
développement
complet à la sortie du cocon. Il contient déjà une
grande quantité de
spermatozoïdes emplissant les testicules et surtout les deux
déférents (Fig.17)
jusqu’au canal commun terminal (Fig.18) De plus, de minuscules glandes
acinoïdes
faisant partie d'un appareil
épigastrique,
sont visibles avant juste en avant du sillon homonyme,
dans sa
lèvre
antérieure (Fig.18).
 |
 |
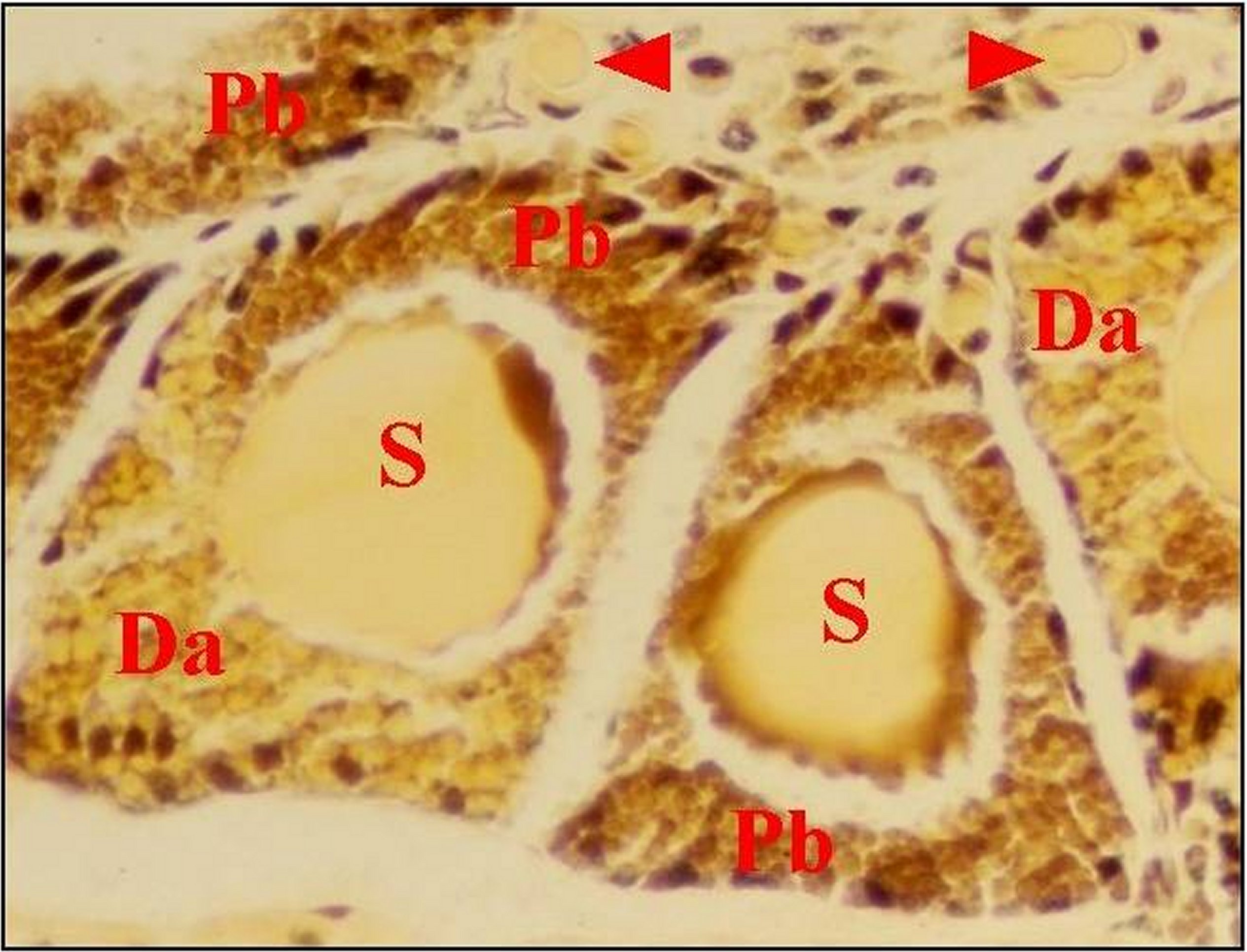
| Fig.17 - Mâle de Mastophora venant de sortir du cocon : Spermatozoïdes murs emplissant le canal déférent. | Fig.18 - Même mâle de Mastophora : spermatozoïdes murs emplissant le canal commun terminal. |
|
A,
appareil épigastrique - Ct, canal
commun terminal - D, diverticule
intestinal - F,
canal déférent -Fe, fente épigastrique - S,
spermatozoïdes murs (©A.Lopez,
C.H.)
|
|
Inconnu
avant les
recherches histologiques de A.Lopez (1985a, 1985b),
il se compose dans le cas de la
femelle des 6 types de glandes séricigènes d' Araneidae dès
le 3eme stade du
développement.
En revanche, l’appareil séricigène
du mâle est réduit à 4
catégories : glandes ampullacées majeures
et
mineures, glandes
piriformes
et glandes aciniformes.
Au
nombre de 4, soit 2 majeures
et 2
mineures, elles ont un corps à deux portions dont
l’épithélium
sécrétant est
formé par 2 catégories d'adénocytes.
Leurs canaux
excréteurs se terminent
respectivement dans les filières
antérieures et moyennes.
Elles
sont assez volumineuses, un peu
irrégulières et «mi-parties» selon
l’expression de Millot car formées par des
cellules proximales et distales à sécrétions
protéiniques se colorant
différemment (Fig.18). Leurs canaux
aboutissent aux
filières antérieures.
 |
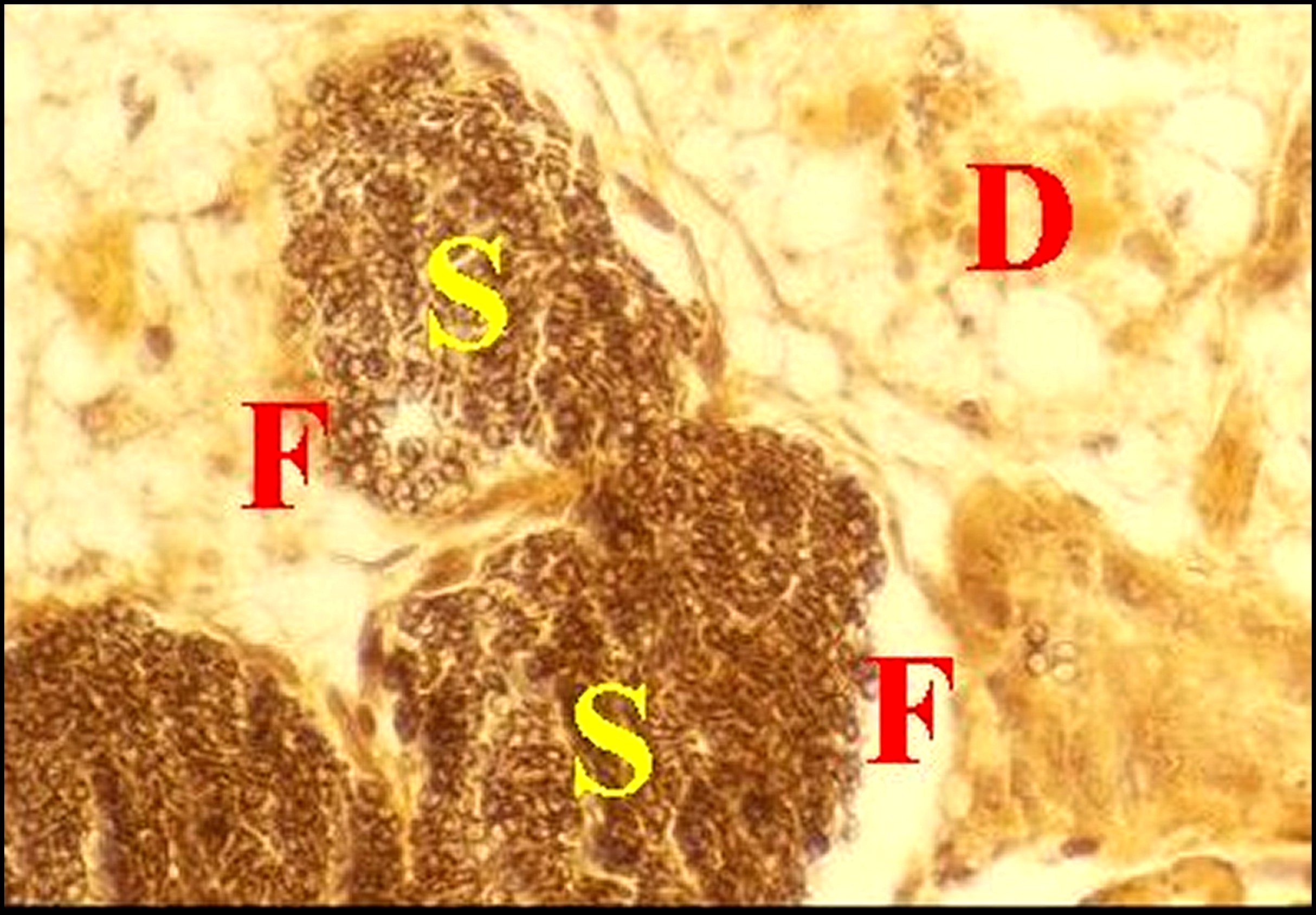
| Fig.19 - Mastophora cornigera, femelle adulte : glandes piriformes. |
| Da,
région distale acidophile - Pb,
région proximale
basophile - S, sécrétion bicolore. Flèches: canaux
excréteurs.
(©A.Lopez,
C.H.)
|
Les
aciniformes de catégorie «B»,
les
moins nombreuses, et de catégorie
«A», se terminent
dans les filières moyennes
et postérieures.
Les
2 paires
d’agrégées sont
remarquables par leur volume
considérable, déjà noté chez les
femelles immatures (Fig.).
Multilobées, elles ont une lumière
anfractueuse et
un aspect arborescent (Fig.20,21). Leur épithélium
simple ne comporte qu’une
seule catégorie d’adénocytes
à
sécrétion glycoprotéinique. Leurs canaux
excréteurs, noduleux et à plusieurs couches
pariétales aboutissent aux filières
postérieures.
Par
ailleurs,
chez Poecilopachys australasia,
autre Cyrtarachnine, les mêmes
glandes ont
un développement
considérable (Fig.37,38). Leurs canaux présentent une paroi à
nodules si étendus
qu'ils
semblent parfois s'en être isolés pour former un tissu
distinct (Fig.).
De
son côté, Celaenia excavata , de même sous-famille, parait
totalement dépourvue de
glandes agrégées,
catégorie dont l'absence est
connue, par ailleurs chez
d'autres Araneidae
construisant pourtant une grande toile non visqueuse
(Cyrtophora)
( Lopez
& Kovoor, 1982).
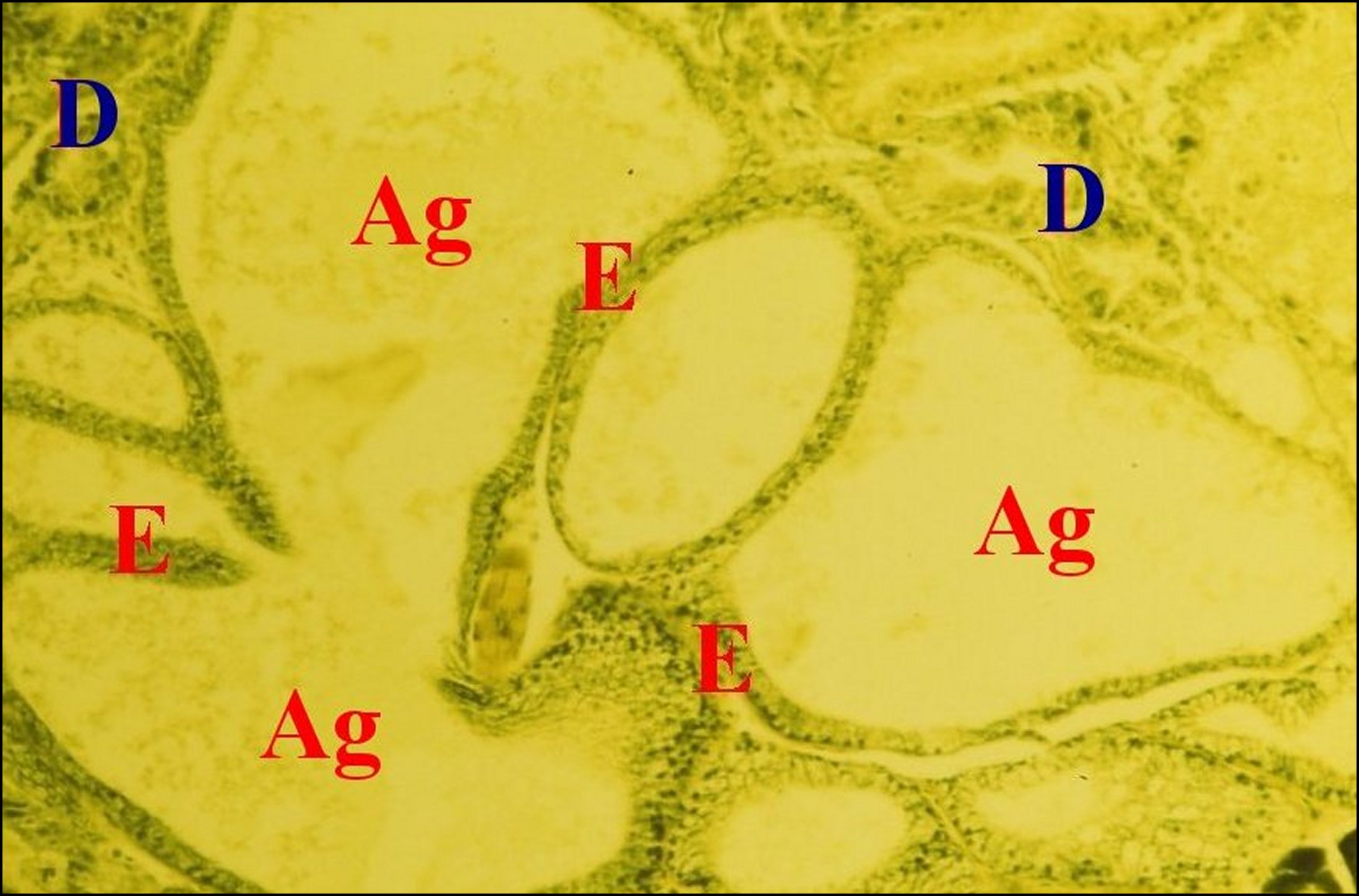 |
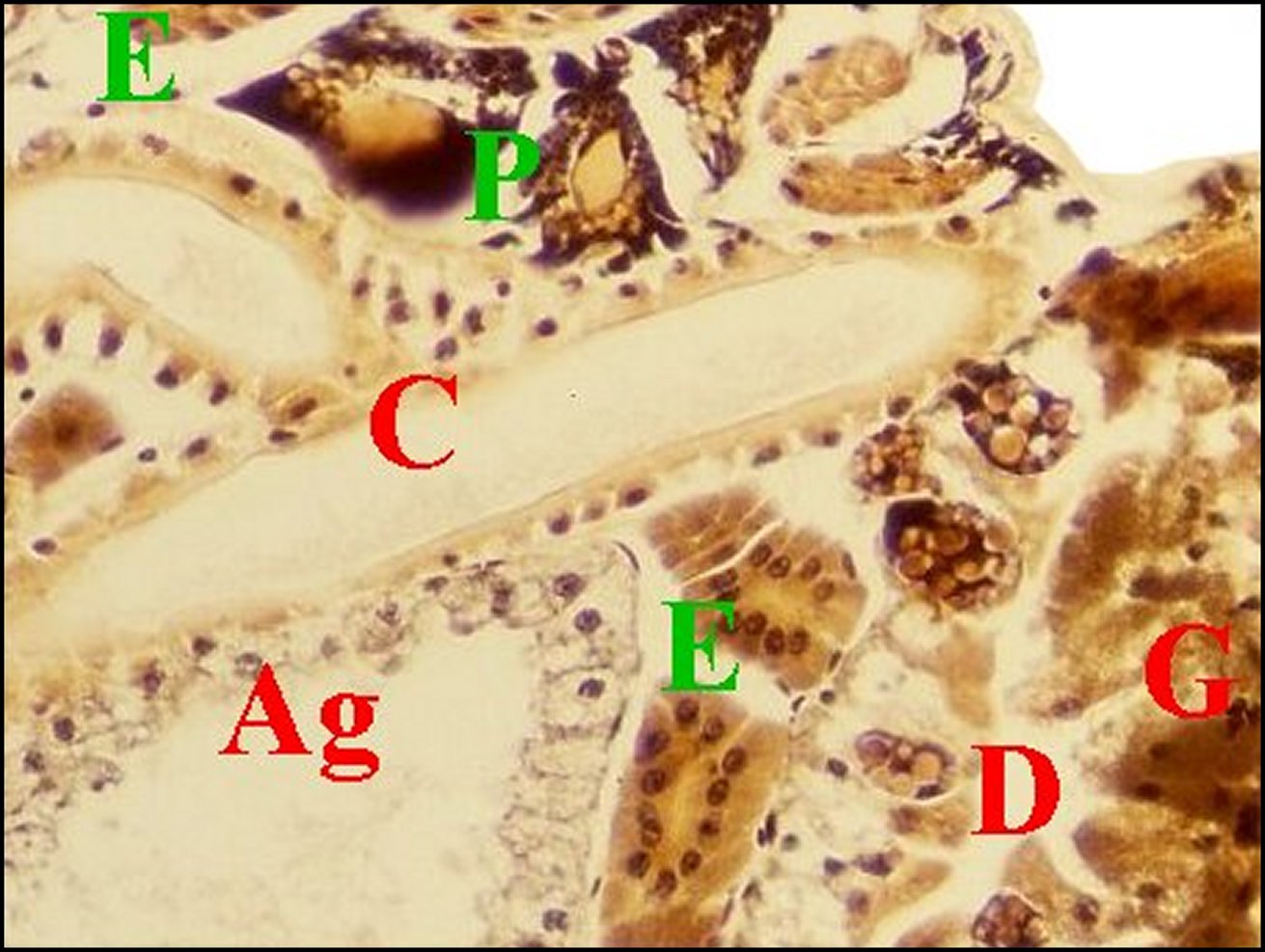
| Fig.20 - Mastophora cornigera, femelle adulte. Vue très partielle de l'abdomen montrant le grand développement des glandes à soie agrégées. |
| Ag,
glandes agrégées - E, leur
épithélium simple - D, diverticules intestinaux (©A.Lopez, C.H.) |
4-7-e - Glandes
flagelliformes
Les
deux glandes flagelliformes
montrent
une partie distale à sécrétion
protéinique et une deuxième proximale
élaborant un produit glycoprotéinique. Leurs canaux
excréteurs sont reçus
également par les filières
postérieures où
chacun d’eux s’associe étroitement
en "triade fonctionnelle" avec une paire de canaux de glandes
agrégées.
Les
six glandes
tubuliformes ont une seule
catégorie de cellules
sécrétoires. Elles se
terminent dans les filières
moyennes et postérieures.
Chez
les Mastophora cornigera
et bisaccata (immatures, femelles adultes,
mâles
mûrs) l'appareil décrit présente
une composition et une structure d’ensemble qui sont typiques des Araneidae, et ne
reflète donc pas la disparition de la
toile orbiculaire. Les glandes flagelliformes produisent
vraisemblablement les
fils axiaux pelotonnés des "bolas" de Mastophora. Le
grand développement des agrégées assure une
sécrétion de "glue" très
abondante.
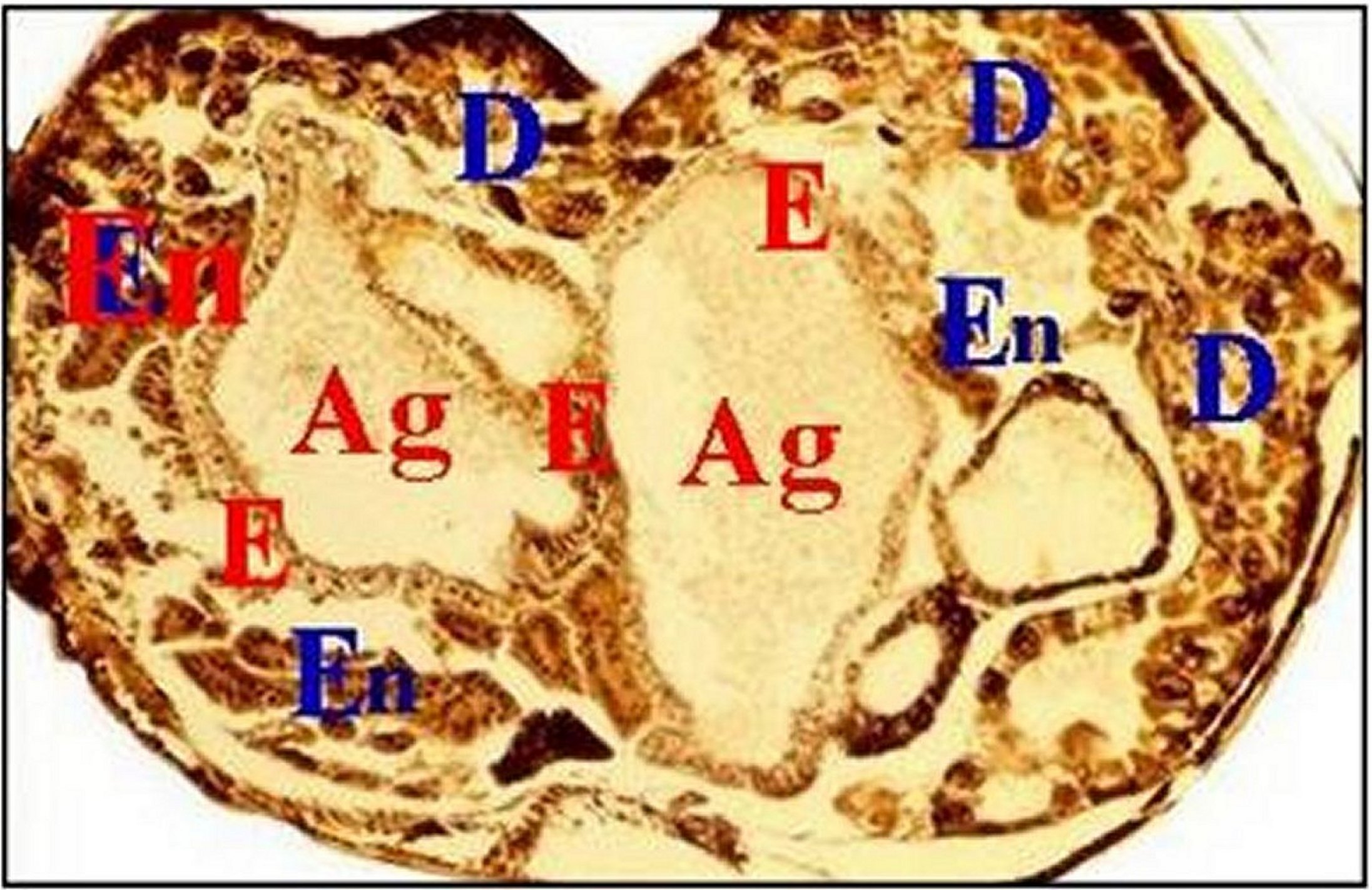 |
| Fig.21 - Mastophora cornigera, femelle immature. Coupe transversale et totale d'abdomen - |
| Ag,
glandes agrégées - E, leur épithélium
simple - D, diverticules intestinaux -
En, tissu endocrinoïde (©A.Lopez,
C.H.)
|
4-8
- Le
tissu folliculaire
endocrinoïde
abdominal
Au
voisinage immédiat
des glandes agrégées et ampullacées des Mastophora ainsi
d'ailleurs que
de Celaenia
et Poecilopachys,
il
existe un tissu très particulier,
d’aspect glandulaire mais
sans canaux excréteurs, entouré
d'hémolymphe,
ne contractant que
des rapports de contiguïté avec l’appareil
séricigène et les
diverticules
chylentériques. Il est réduit chez le mâle
de Mastophora (Fig.22,23),
bien plus développé chez les femelles immatures du
même genre (Fig.24,25) et
atteint un volume considérable lorsqu’elles deviennent adultes
(Fig.26 à 28).
Dans
tous les cas,
ses cellules sont bien
individualisées, de tailles et de forme
assez constante
globuleuse, polyédrique, cuboïdale ou même conique.
Leur cytoplasme est
acidophile, finement grenu, parfois criblé de très
petites vacuoles. Leur noyau
est arrondi, clair, nucléolé, riche en chromatine bien
visible. Elles se
réunissent en petits massifs et cordons peu sinueux, arrondis
dans les coupes
transversales, compacts ou montrant souvent une cavité axiale
très étroite,
presque virtuelle et qui n’est pas une vraie lumière (Fig.23).
Ces groupements
de cellules sont assez peu nombreux, bien isolés les uns
des autres, de
taille relativement grande chez les immatures (Fig.23,24), en
quantité beaucoup
plus grande, tassés les uns contre les autres et plus petits
chez les femelles
adultes (Fig.25 à 27).
Ils
se présentent
comme des structures sacciformes collabées,
entièrement
closes, dépourvues de
toute structure canalaire, baignées directement par
l’hémolymphe. Un tel
ensemble de caractères est très évocateur de leur
nature «endocrinienne» et
n’est pas sans évoquer les follicules (vésicules)
thyroidiens de
Vertébrés. Faute d’une meilleure appellation, le tissu
peut donc être qualifié,
selon Lopez (1998),
de
"folliculaire endocrinoïde". Il
s’insinue entre les organes voisins, notamment les lobes des
agrégées et les
diverticules chylentériques
(Fig.24) qui semblent d’ailleurs
les
déformer par
compression. De plus, il tend à se répartir en quatre
amas : deux
ventraux, paramédians, situés un peu en arrière du
niveau des phyllotrachées
(poumons) et deux
latéro-dorsaux, très externes. Les amas sont
seulement accolés aux
glandes séricigènes
agrégées et entourent
l’origine de leurs canaux
excréteurs
(Fig.24, 26), sans solution de continuité. Le tissu folliculaire
endocrinoïde
du mâle de Mastophora
cornigera se limite à deux petits massifs cellulaires
symétriques
pouvant montrer quelques signes de
dégénérescence : condensation ou
vacuolisation exagérées du cytoplasme, pycnose
nucléaire (Fig.22,23)
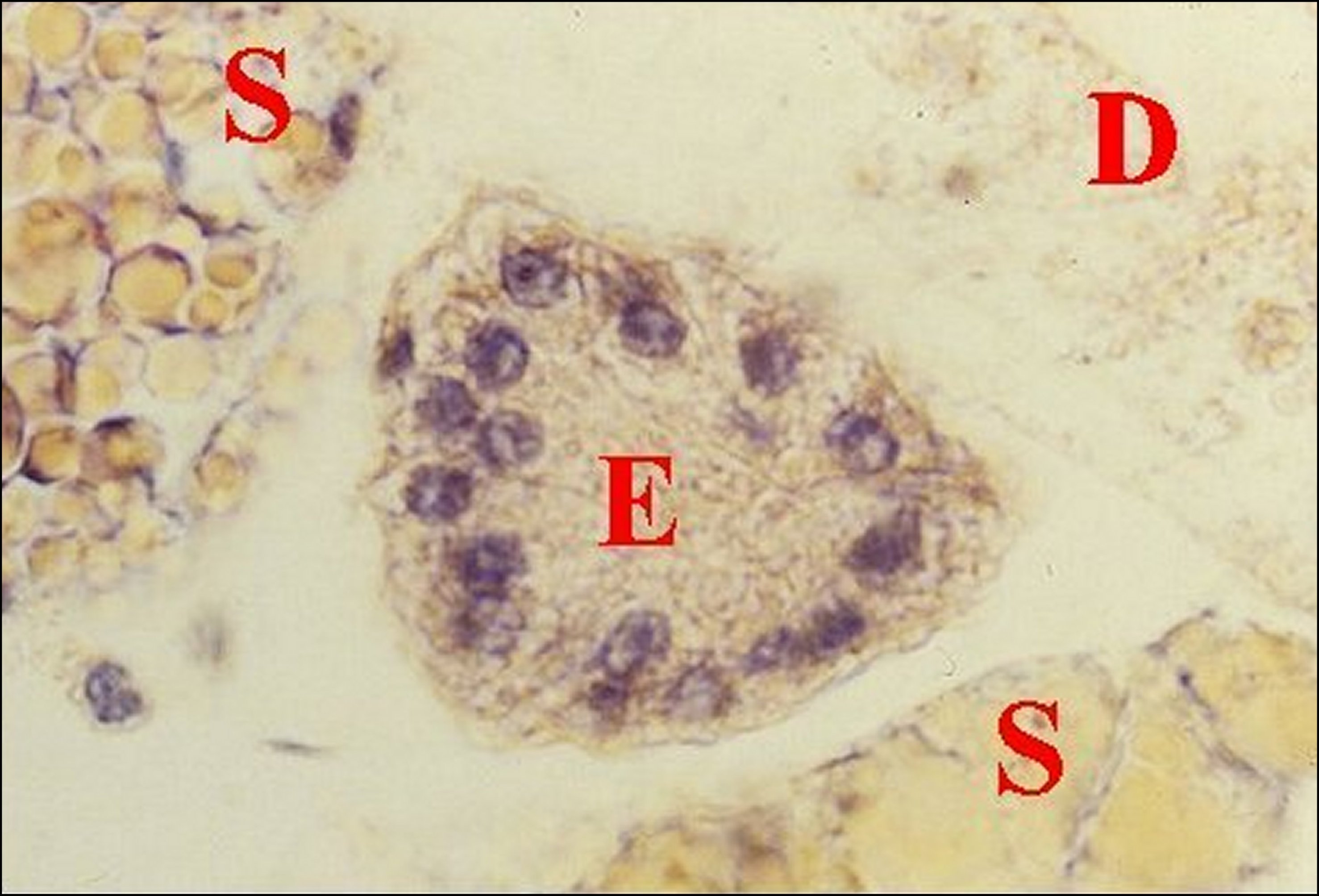 |
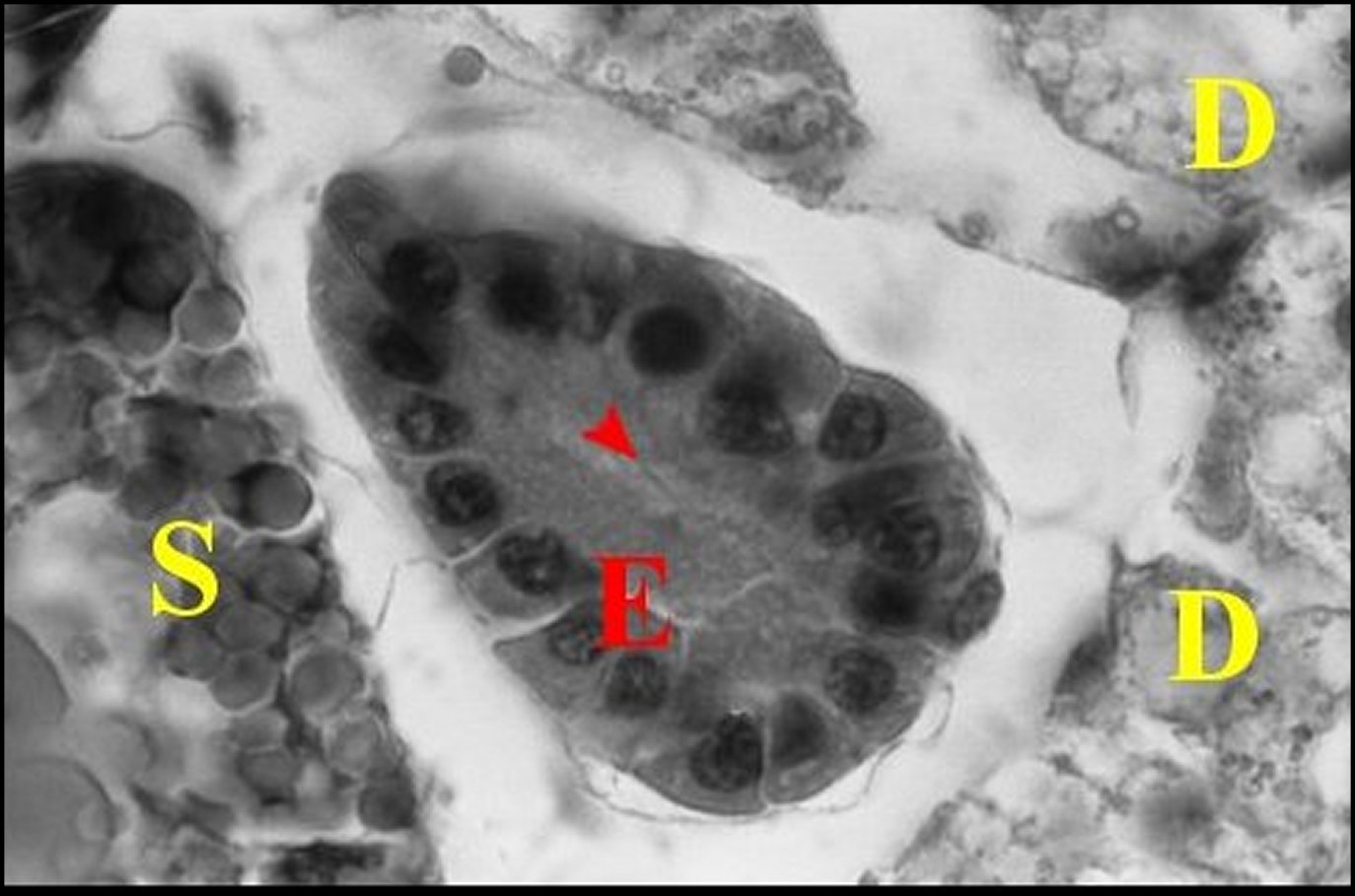 |
 |
 |
|
Fig.22
- Mastophora
cornigera, mâle :
ilôt
de tissu
folliculaire endocrinoïde.
|
Fig.23
- Mastophora cornigera, même
mâle :
autre ilôt
de tissu folliculaire endocrinoïde (photo noir et blanc) |
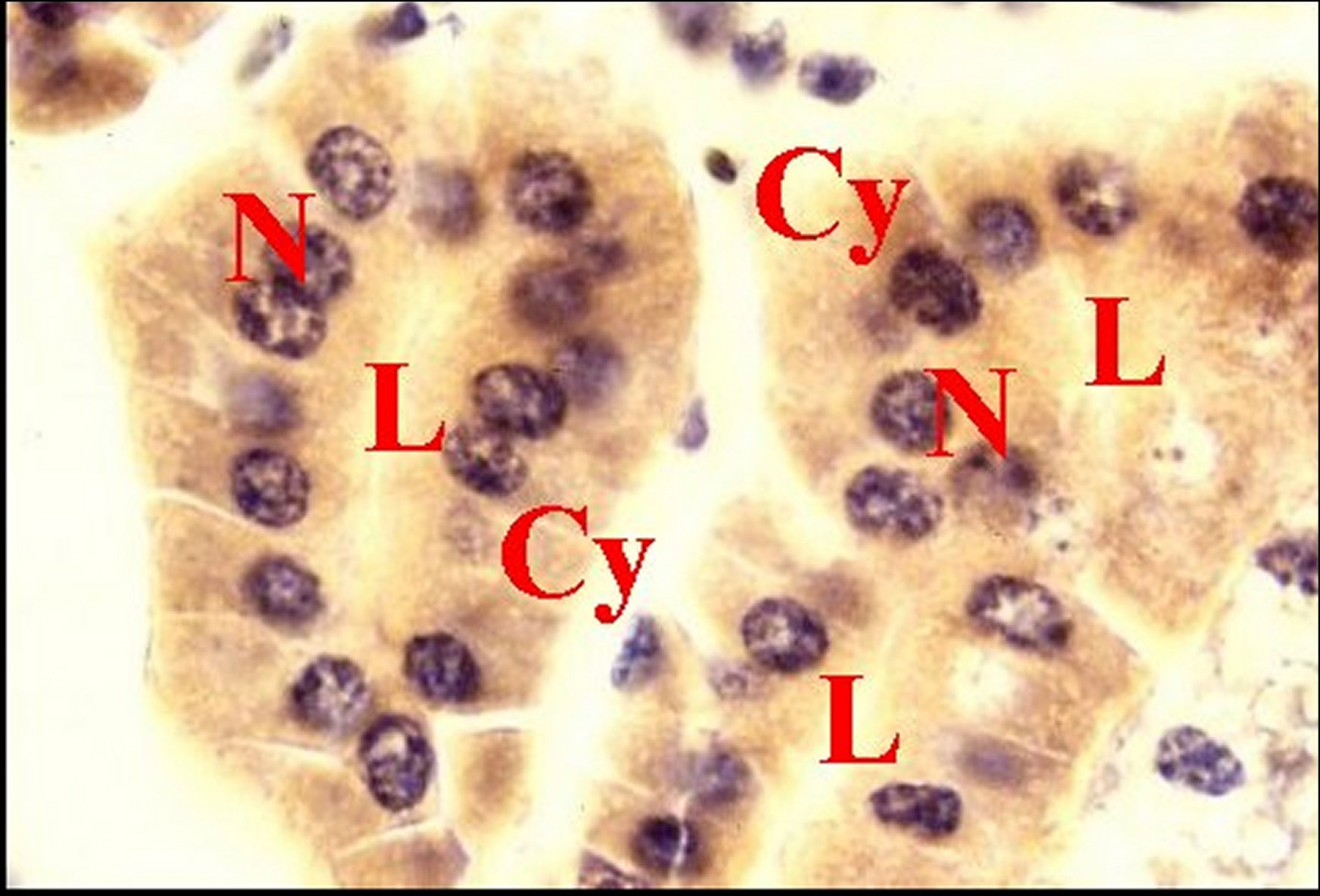
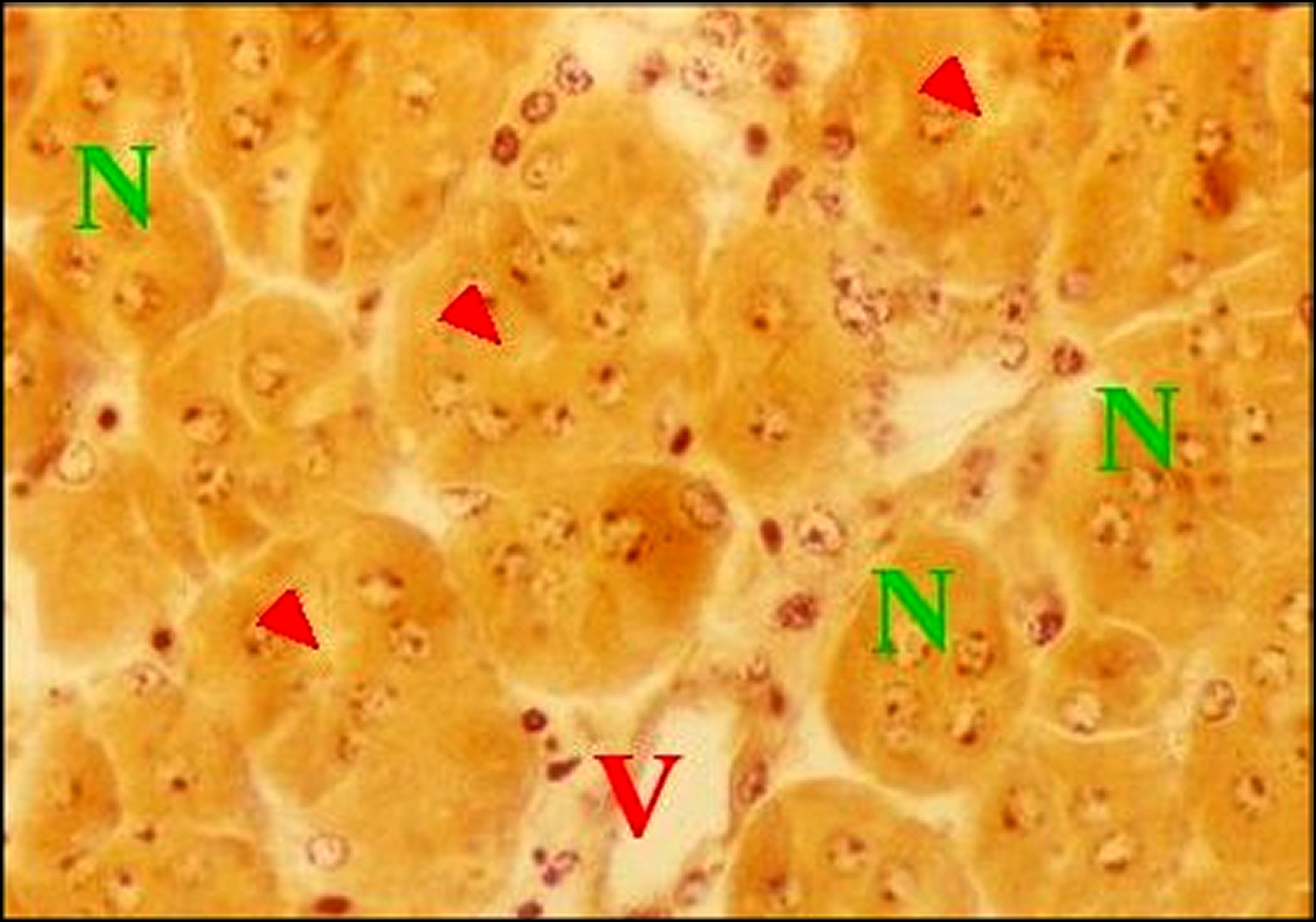
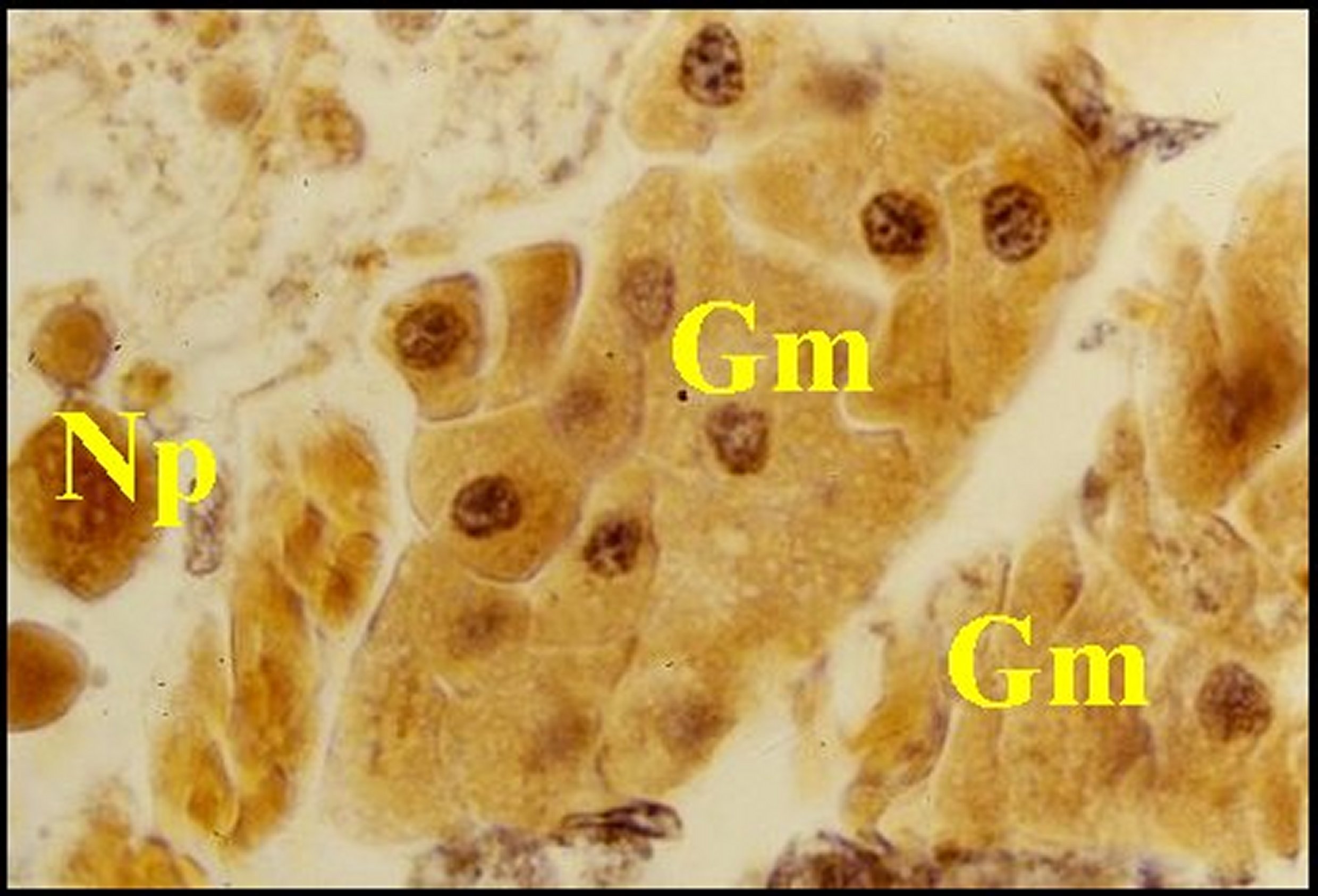
Fig.24 - Mastophora cornigera, femelle immature : Emplacement du tissu endocrinoïde dans une vue partielle d'abdomen. | Fig.25 - Mastophora cornigera, femelle immature, tissu endocrinoïde. |
| C, canal de glande agrégée - Cy, cytoplasmes de cellules endocrinoïdes - E, tissu endocrinoïde - D, diverticule intestinal -G, cellules à guanine - L, cavité ("lumière") centrale - N, noyaux de cellulesendocrinoïdes S, glande à soie - Flèche : cavité centrale (©A.Lopez, C.H.). | |||
 |
 |
 |
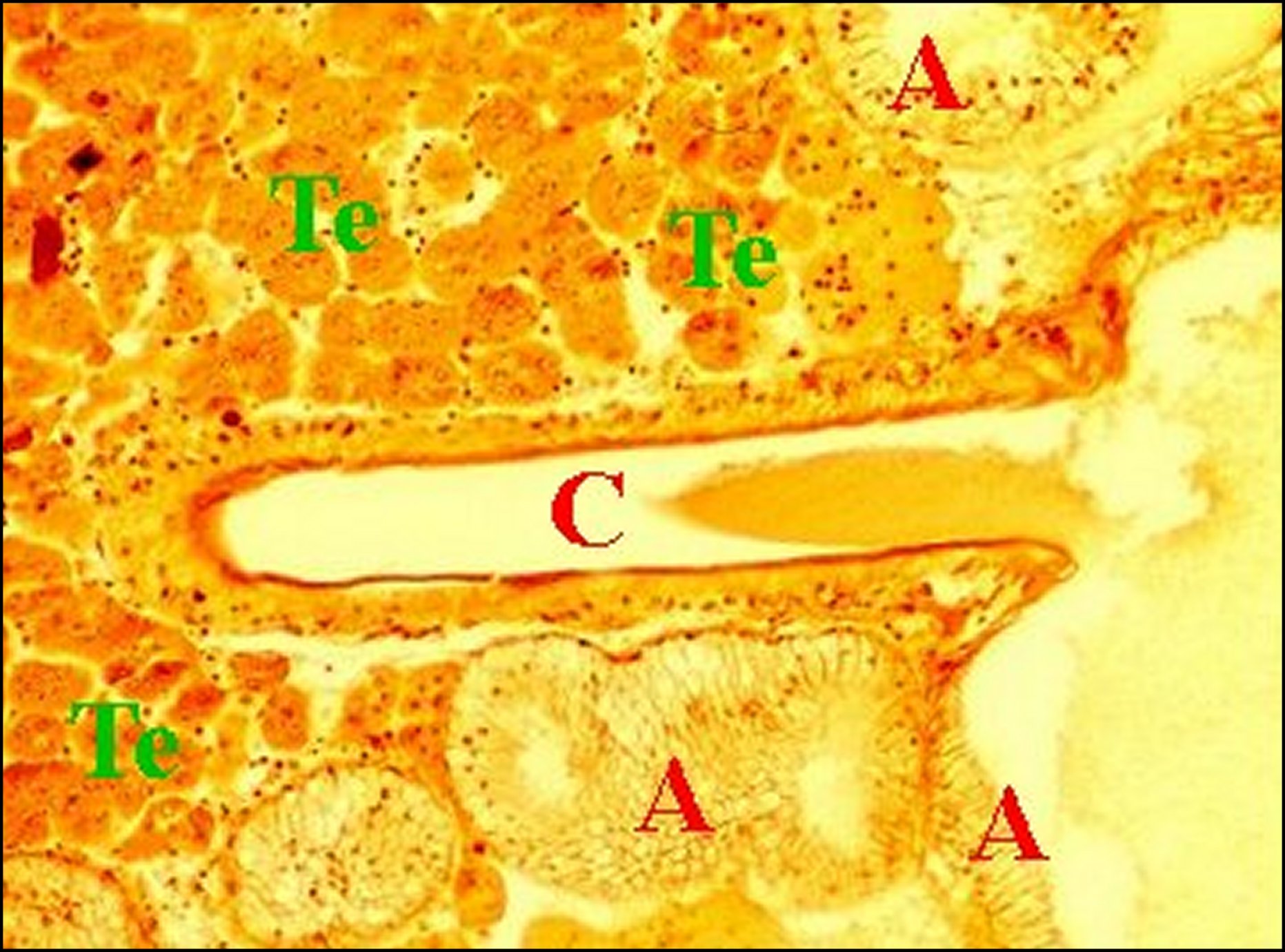
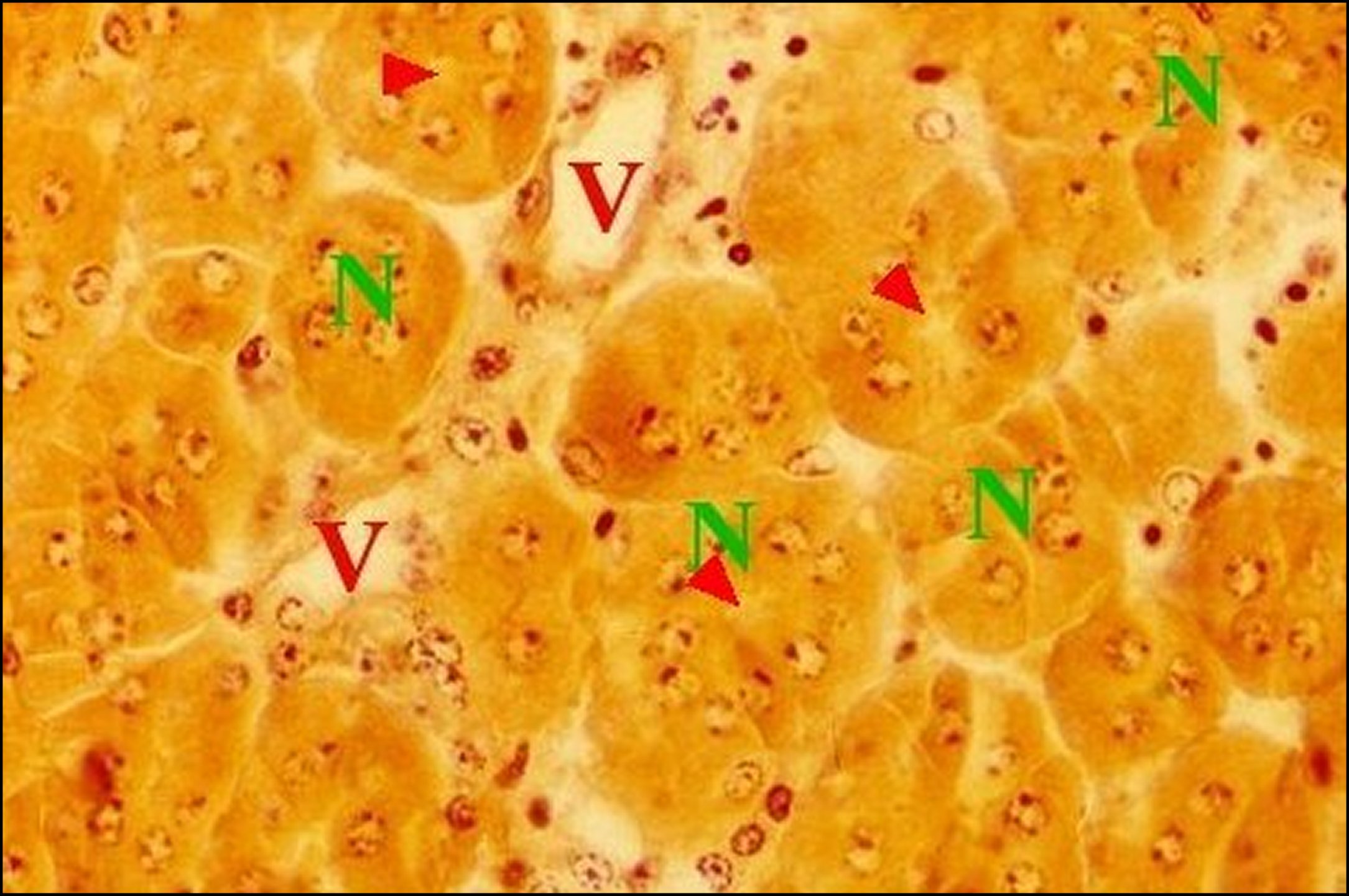
| Fig.26 - Mastophora cornigera femelle adulte : vue trés partielle d'une coupe d'abdomen. | Fig.27 - Mastophora cornigera, femelle adulte.Détails (cellules) du tissu endocrinoïde. | Fig. 28 - Mastophora cornigera, femelle adulte. Autres détails (cellules) du tissu endocrinoïde. |
| A, corps de glandes agrégées - C, canal excréteur - N, noyaux des cellules endocrinoïdes disposées en ilôts juxtaposés - Te, tissu endocrinoïde folliculaire - V, vaisseaux. Les flèches désignent des cavités (©A.Lopez, C.H.) | ||
Elle
n'a été
étudiée (sommairement) par Lopez que chez Mastophora
bisaccata.
Les
amas cellulaires sont entourés par une lame
basale mince formée d’un matériel finement
granuleux
(Fig.30). Il semble exister
une discrète polarité cellulaire, notamment dans les amas
« évidés »
en leur centre car le plasmalemme
tend
à former des invaginations intracytoplasmiques au-dessus
de
la lame basale.
Le noyau,
clair et arrondi,
renferme de la chromatine
dispersée
dans son nucléoplasme et
formant aussi de
grosses mottes marginales. Le réticulum
endoplasmique est
remarquablement développé, lisse, et se
compose de cisternae
pouvant se dilater
en chapelets vésiculaire. Les autres organites
sub-cellulaires
sont des mitochondries allongées
pouvant présenter des crêtes
tubulaires, des dictyosomes,
de petits lysosomes, des polysomes
et des microtubules épars (Fig.29,30).
 |
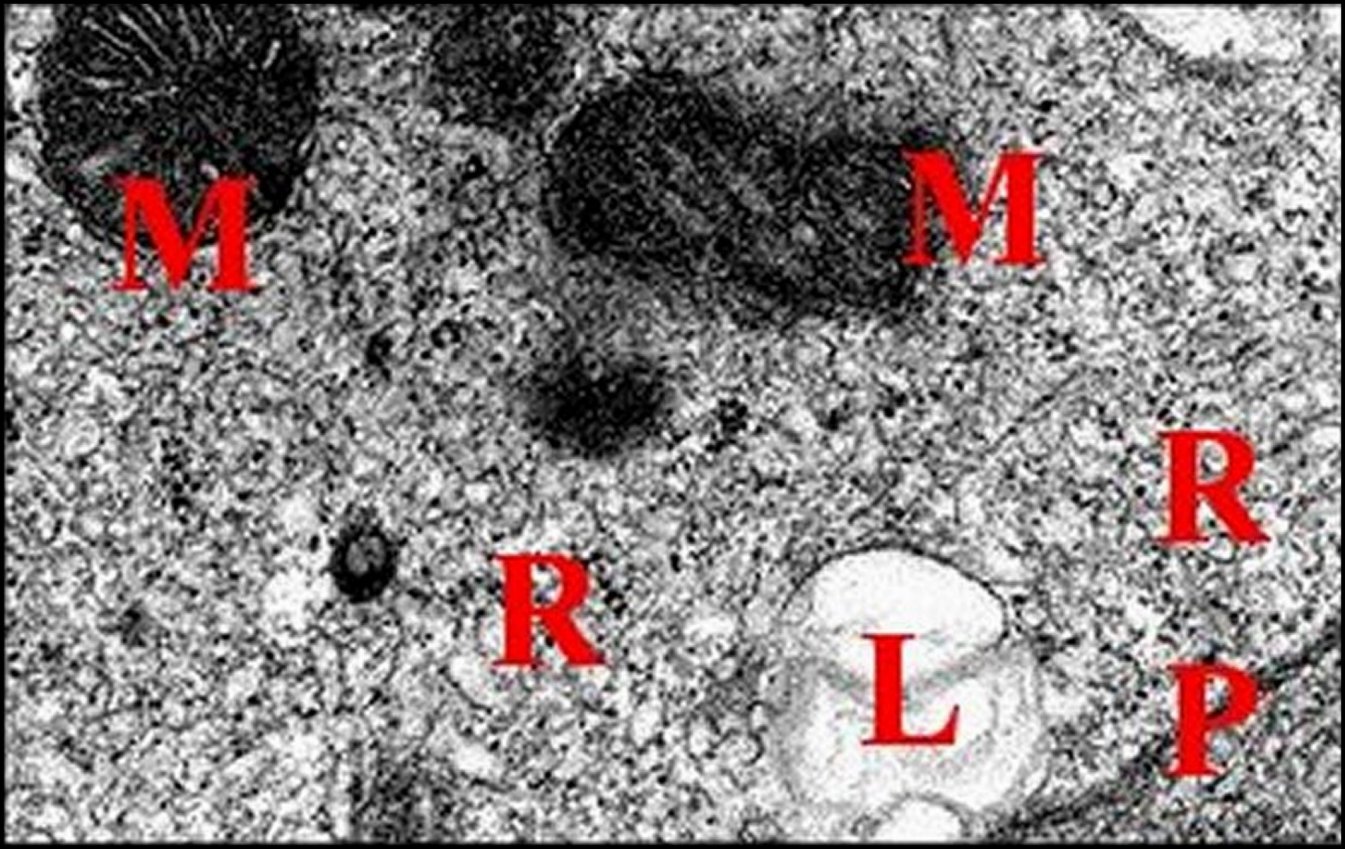 |
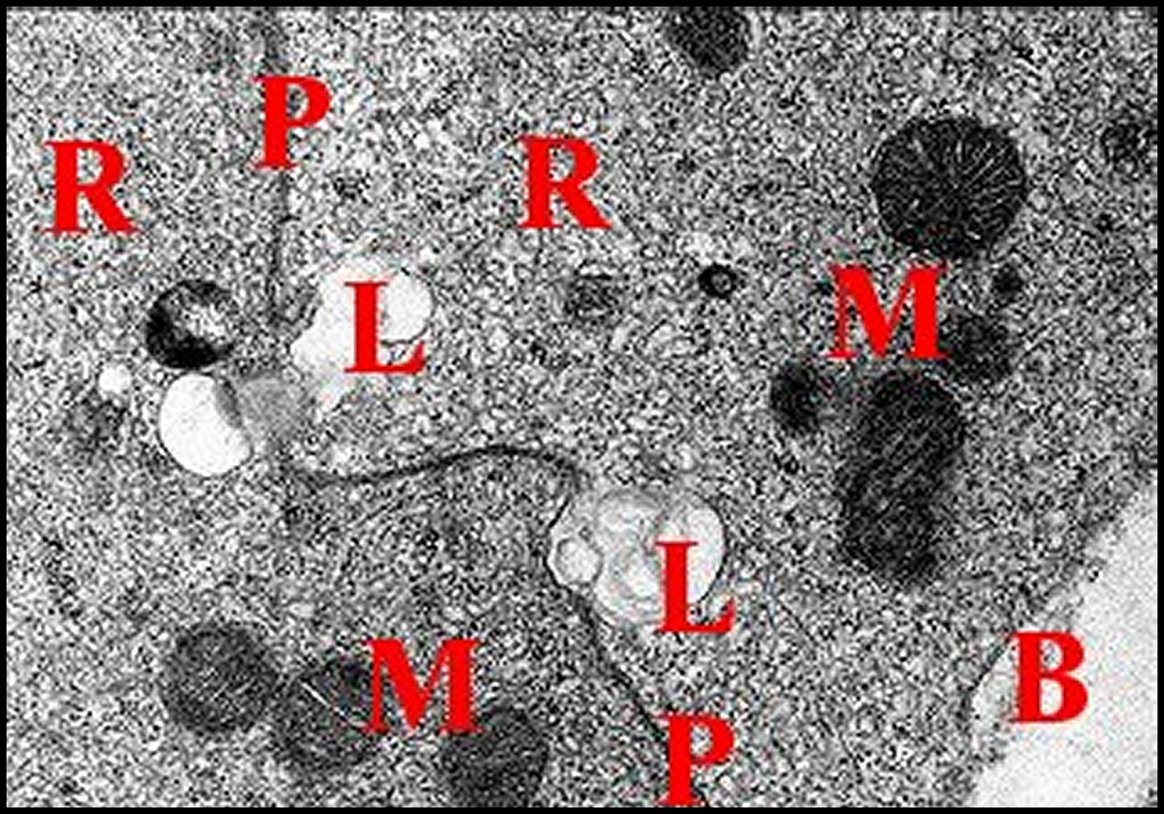
| Fig.29 - Mastophora bisaccata femelle, cellules endocrinoïdes: organites subcellulaires | Fig.30 - Mastophora, la même, tissu endocrinoïde : détails des organites |
| B, lame basale - L, lysosomes - M, mitochondries à crêtes tubulaires - P, plasmalemme - R, réticulum lisse très développé (©A.Lopez, M.E.T). | |
5-1 - Considérations anatomo-physiologiques
L'appareil séricigène de Mastophora
(immatures,
femelles adultes, mâles mûrs) a une composition et une
structure d’ensemble qui
sont typiques des Araneidae,
et ne reflète donc pas la
disparition de la toile orbiculaire.
Les glandes flagelliformes
produisent
vraisemblablement les fils axiaux pelotonnés des "bolas" tandis
que
le grand développement des agrégées
(Fig.20 et 21)
assure une
sécrétion de
"glue" très abondante, ce qui garantit la production
répétée des
globules, comme chez Poecilopachys -
pourvue aussi
d'énormes glandes de la même
catégorie (Fig.37,38) - et la
mise en place des enduits
visqueux centraux sur les « fils en pont » de la
fausse spirale. Les glandes tubuliformes
élaborent la soie des cocons ovigères globuleux à paroi rugueuse et
parcheminée, pédiculisés et pourvus parfois
d'un socle les
fixant sur un réseau de quelques fils tendu dans la
végétation (Fig. 31)
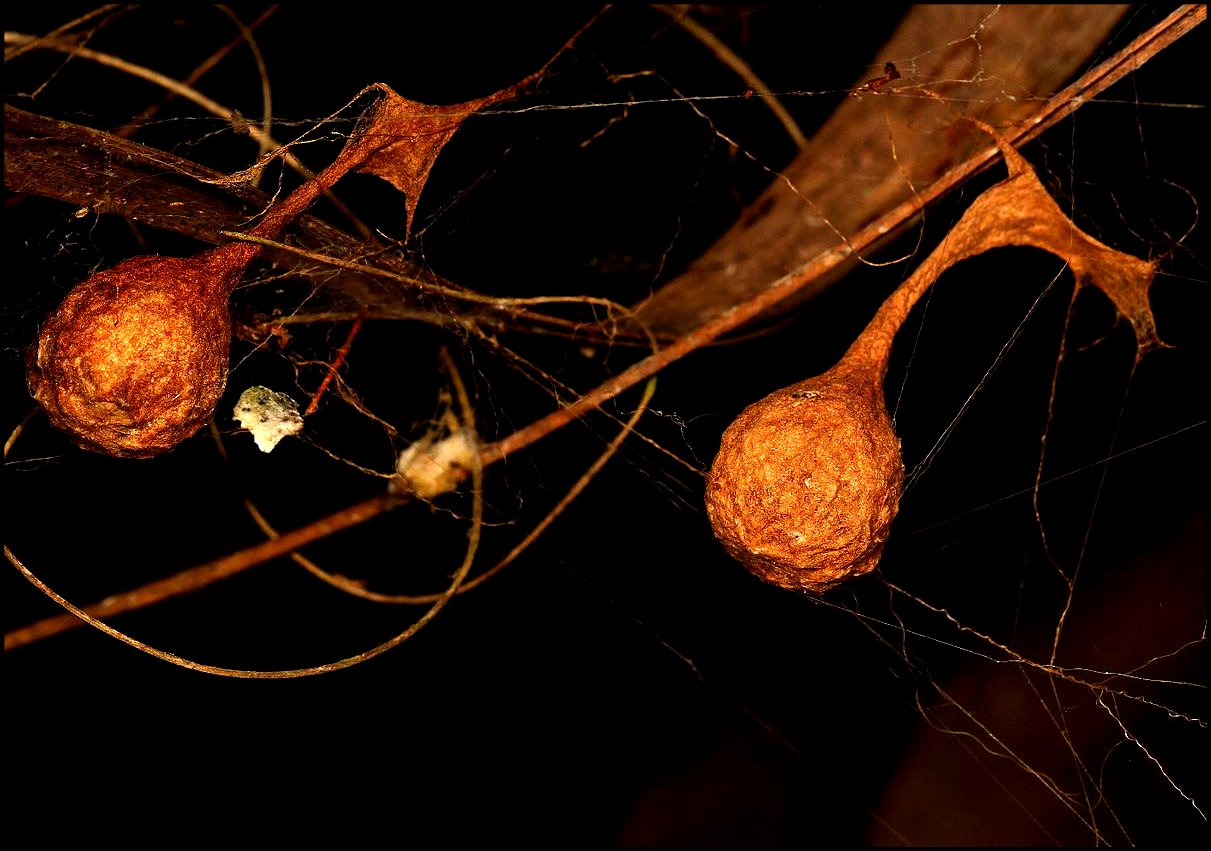 |
| Fig. 31 - Cocons de Mastophora cornigera (?) |
En
revanche,
dans le même cadre des Araignées
à allomones, l'appareil séricigène de Celaenia semble
bien être dépourvu de glandes
agrégées (Fig.35,36),
ce qui explique un comportement de chasse excluant tout fil de
capture. Dans aucun de ces cas, les agrégées,
lorsqu’elles existent, et
toutes les autres catégories glandulaires ne comportent aucune
cellule spéciale
qui puisse être comparée à celles des glandes
botryoïdes chez Kaira
alba (Lopez,1986).
Les
diverticules
intestinaux ou chylentériques
pénétrant dans les
cornes du prosoma où
Lévi (2003)
rappelle
leur présence d'après Lopez (mais pas celle de
l'endocrinoïde !), sont
largement ouverts
dans le thoracenteron (Fig. 8
à 12,32,33), sans
toutefois s'engager
dans les excroissances cornues
latérales (Fig.12). Il s'
ensuit que les Mastophora font
partie du groupe des Araneidae où
cette partie toute
antérieure et céphalothoracique de l'intestin est
remarquablement développée
(Millot,1931).
Les
diverticules
pourraient être impliqués dans deux manoeuvres
particulières
qu'effectuent
les Mastophora (Hutchinson,1903 ; Eberhard,1980) ;
l'absorption des bolas
inutilisés ou défectueux que
l'araignée ingère afin de
maintenir sa réserve hydroprotéinique ; la
régurgitation d'un liquide
malodorant qu' elle émet lorsqu'on l'importune et
réabsorbe quand cesse la
perturbation. Ledit liquide pourrait bien être le contenu des
diverticules
mobilisés par la contraction des muscles prosomatiques adjacents
et sans rapport évident avec les guaninocytes voisins (Fig. 32,33).
5-1-d - Tissu
folliculaire endocrinoïde
5-1-d-a - Rapports
avec la glande de mue
Le
tissu folliculaire
endocrinoïde abdominal est
bien différent du tissu
interstitiel adipoïde
étroitement associé avec l’intestin (Millot, 1926), ne
peut correspondre à des
glandes séricigènes
ébauchées ou
involuées et paraît aujourd’hui ne plus
pouvoir être considéré comme un simple contingent
postérieur de la glande de
mue (Fig.34,35), première conception erronée de
Lopez
(1985a)
chez Mastophora,
peut être inspirée
par celui des Mygalomorphes.
Découverte
et décrite
par Millot (1930) sous
le nom de "tissu endocrine", cette dernière présente dans
tout
l’ordre des Araignées une stricte localisation
céphalothoracique (Streble,
1966 ; Bonaric, 1980)
à
l’exception toutefois de certaines Mygales, dont les Liphistiidae,
où on la retrouve jusque dans l’abdomen au
contact des gros nerfs (Legendre, 1981 ; Legendre & Lopez,1981).
Son rôle dans les mécanismes de l’ecdysis a
été évoqué
par Legendre (1959) qui la considère comme un homologue
mésodermique de celle
des Insectes, et confirmé par Bonaric (1980) qui
lui a attribué son nom définitif.
Cette
glande de mue
(Fig.34,35) adopte dans le prosoma une disposition spatiale
qui
est en
grande
partie métamérique («organe antérieur»
et «organe postérieur», «groupes
latéraux») alors que le tissu folliculaire endocrinoide
abdominal ne semble pas
en présenter une. De
plus, des examens comparatifs montrent que
leurs aspects
histologiques ne sont pas exactement superposables. Contrairement au
tissu folliculaire
de Mastophora (Fig.34), Poecilopachys (Fig.35) et
les autres Araignées, les cellules de la glande de mue ont une
taille assez
variable et une forme plus irrégulière. Elles se groupent
plutôt en rubans et
en travées compactes où il n’y a pas de
«lumière» ; leur association avec
des néphrocytes et
des hémocytes est
quasi constante
(Fig.34). Les
caractères ultrastructuraux sont en revanche assez voisins (réticulum endoplasmique lisse développé,
crêtes mitochondriales
tubuleuses).
 |
 |
(Céphalothorax, vue trés partielle). |
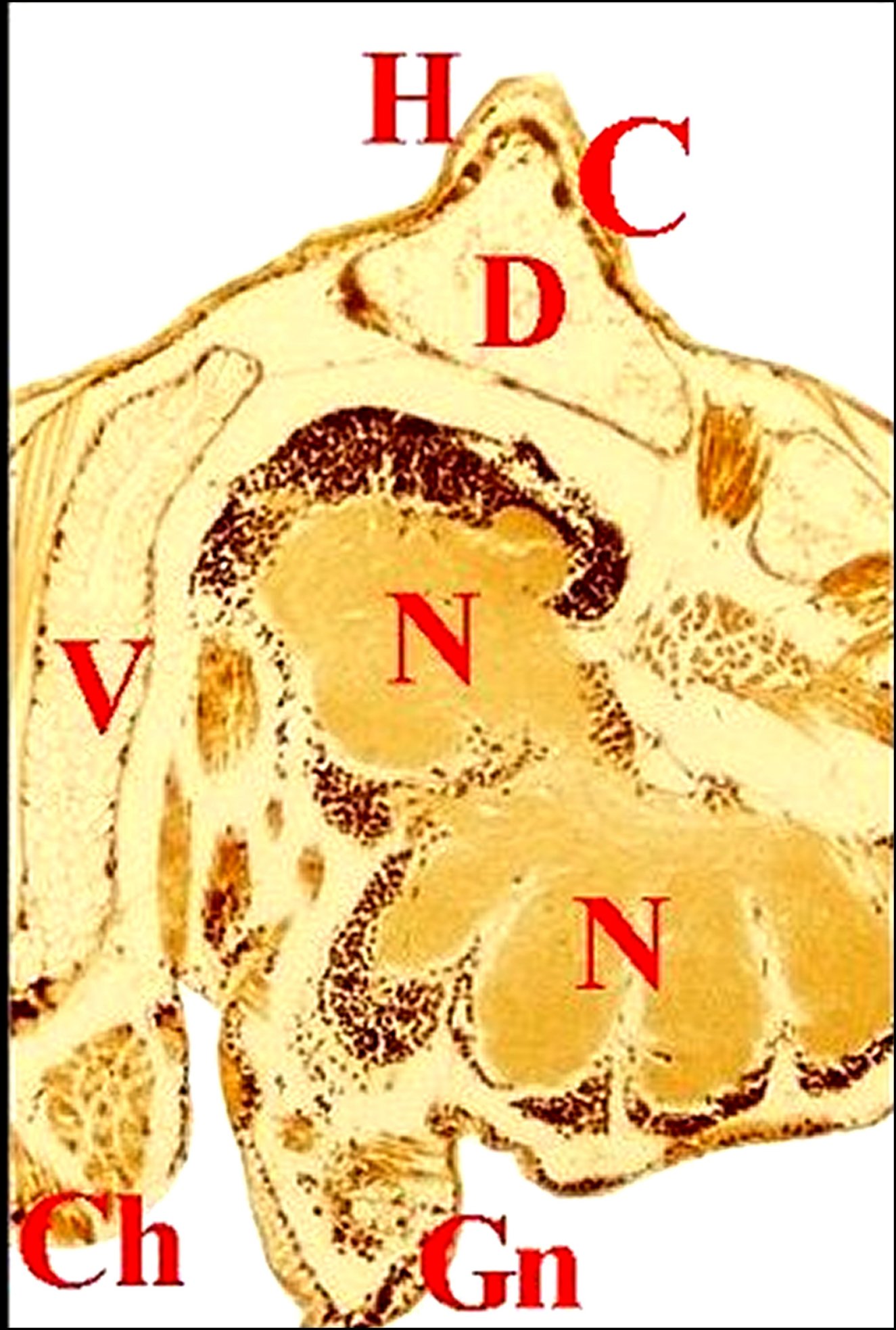
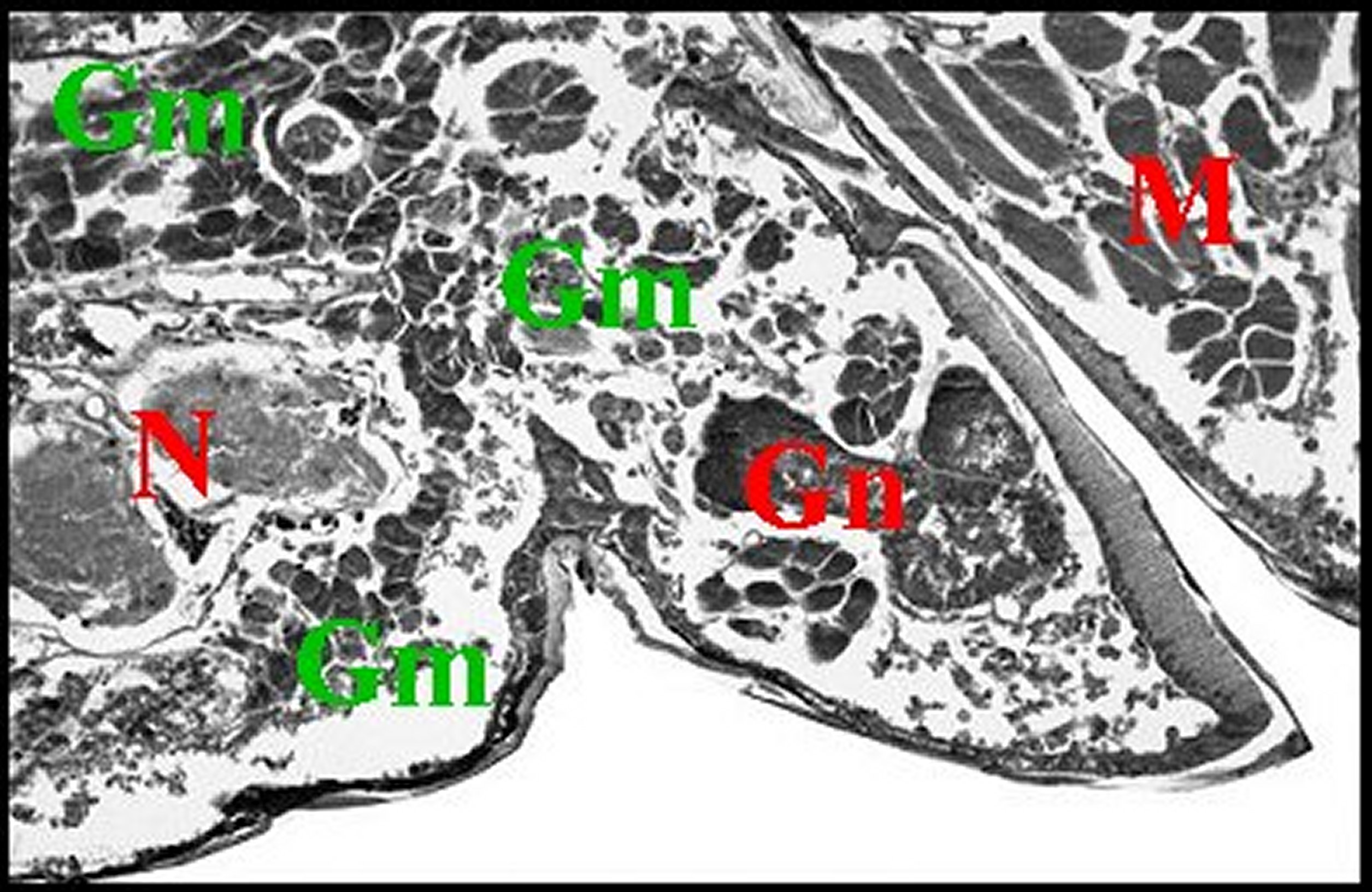
Fig.35- Poecilopachys
australasia femelle, glande de mue (Céphalothorax en coupe parasagittale). |
| Gm,
glande de mue prosomatique - Gn,
gnathocoxe avec ses glandes
"salivaires" - M, muscles - N,
ganglions nerveux sous-oesophagiens- Np, néphrocytes (©A.Lopez, C.H.). |
|
Sur
le plan fonctionnel, il est
très
possible que le tissu folliculaire endocrinoïde abdominal soit
impliqué dans
l’attraction et la capture de proies, le produit des
régurgitations et
l'’épiderme des pattes étant à exclure, du moins
chez les femelles adultes. Ses
relations avec la substance attractive de nature sémiochimique
indiscutable autorisent deux
hypothèses, la
plus séduisante étant inspirée dans le cadre des Araignées à
allomones par le
mode d’action des ecdystéroïdes, dont on sait qu’ils sont
responsables du
déterminisme de la mue et de autres processus mettant en jeu la
différenciation
cellulaire (métamorphoses, formation des œufs) aussi bien chez
les Insectes que
chez les Araignées. Elle
est étayée, dans le cas des Mastophora
et autres Cyrtarachninae par les
observations d’auteurs
nord-américains
tels qu’ Eberhard (1977) privilégiant
le corps même de
l’Araignée et excluant formellement une quelconque attraction
par les produits
des glandes à soie,
ce qui confirme la première
hypothèse de Lopez.
Il s
'agit là d'un exemple type de
mimétisme chimique agressif.
5-2 -
Considérations
phylogéniques
Ainsi qu'évoqué au début de ses observations sur les «Araignées excentriques» du nouveau Monde par Lopez (1985) et confirmé ensuite ultérieurement (Lopez, 1998), il existe une ressemblance certaine entre les Mastophora, Celaenia et Poecilopachys non seulement par leur équipement séricigène (composante agrégée«arborescente» en particulier) mais surtout, par le tissu folliculaire endocrinoïde abdominal dont elles sont pourvues, particularité majeure ignorée de Levi (2003), tout comme les glandes botryoïdes de Kaira par ce même auteur (Levi,1993).
 |
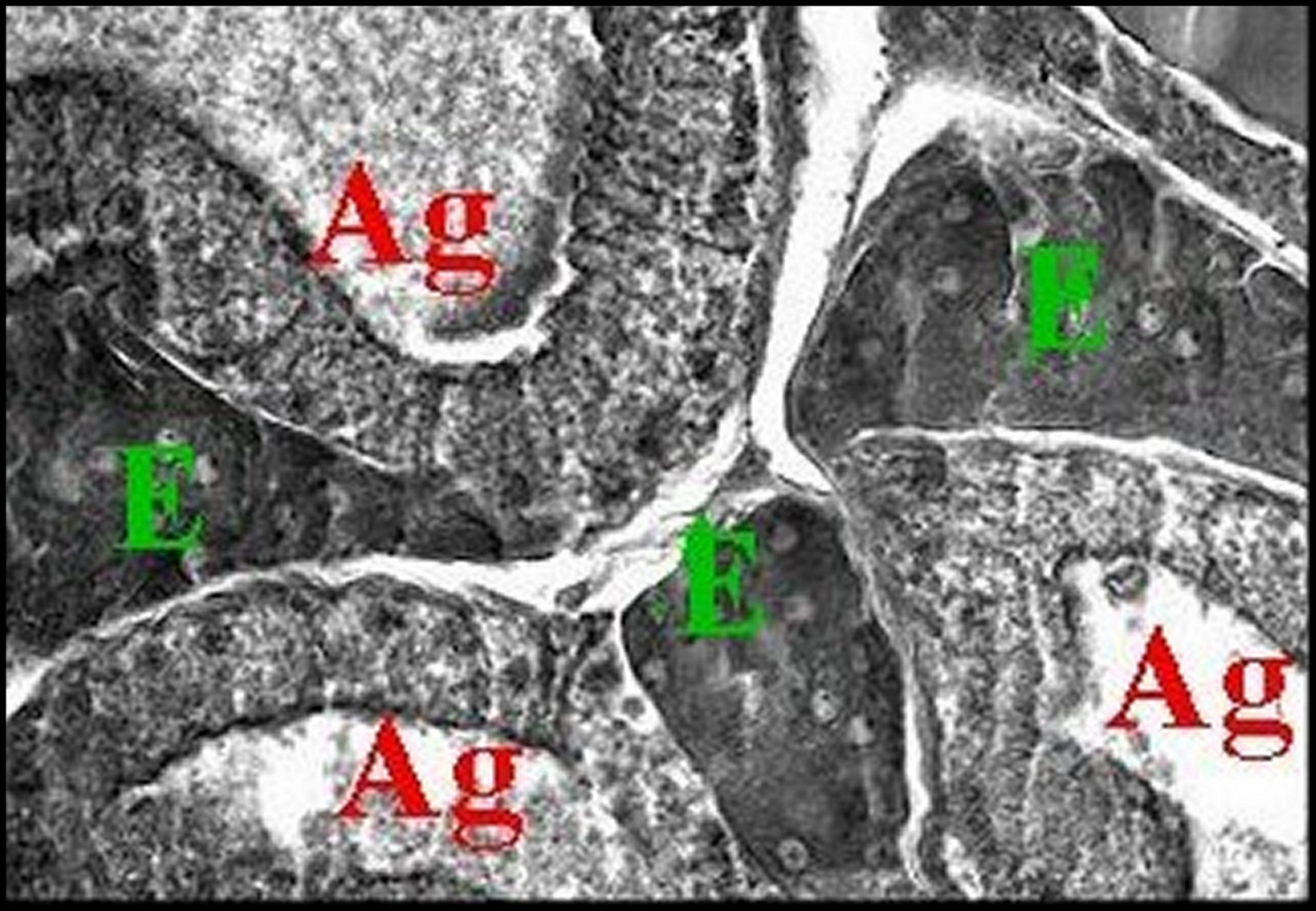 |
| Fig.36- Celaenia
excavata, femelle : ilôt de tissu endocrinoïde,
détail Les glandes agrégées sont absentes.
Tubuliformes visibles. |
Fig.37
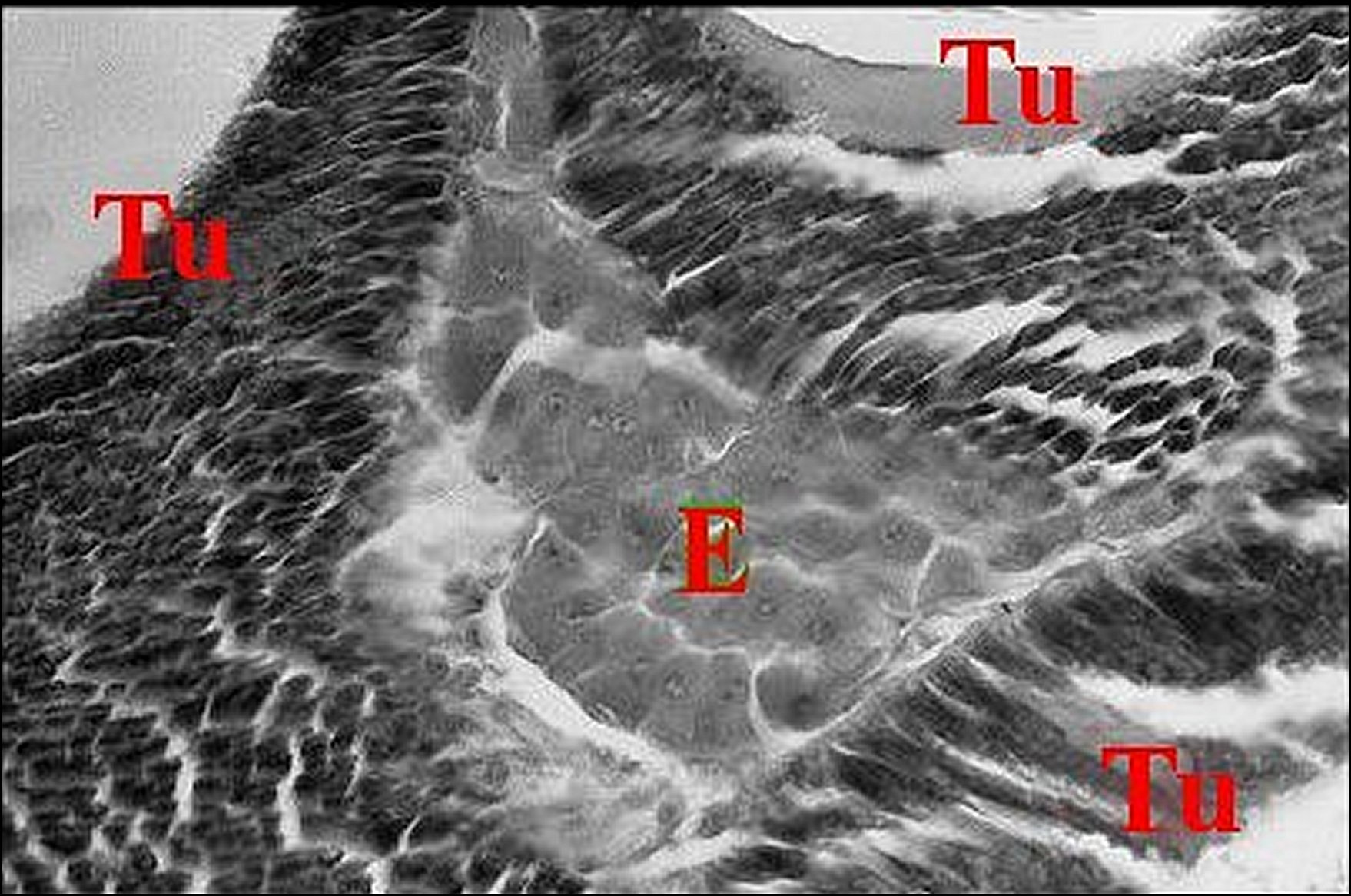
-Poecilopachys australasia, femelle : trois ilôts de
tissu endocrinoïde et glandes agrégées énormes |
| Ag, glandes à soie agrégées - E,tissu endocrinoïde - Tu, glandes à soie tubuliformes (©A.Lopez, C.H.). | |
Eberhard (1980)
dans
son étude sur Mastophora dizzydeani et
Robinson (1982), dans sa monographie sur
les Araignées de
Papouasie, avaient
proposé de rattacher les Mastophoreae,
les Celaenieae et les
Cyrtarachneae,
dont Poecilopachys,
à un même groupe
monophylétique.Leurs arguments, repris par Stowe (1986)
n’étaient pas
anatomiques mais uniquement d’ordre éthologique :
prédation presque
exclusive sur des Hétérocères, du moins chez les
Mastophorinae ;
homologie
des fils «en ponts» («spanning-threads») et des
«bolas», peignés avec les pattes
IV lors du tissage, tous pourvus d’une glue adhésive pour
les
Lépidoptères ; régurgitations malodorantes et
peut être
répugnatoires ; flexion particulière des pattes
I et II lors du
repos.
Ces
particularités
disparates peuvent bien être considérées comme
des synapomorphies mais
paraissent néanmoins plus accessoires
que les caractères anatomiques internes, presque toujours
ignorés ou au mieux
négligés. La structure fondamentale de l’appareil
séricigène reste typique
des Araneidae et
n’est donc pas un argument formel. En
revanche, le tissu folliculaire
endocrinoïde se présente
comme une
caractéristique majeure, commune aux trois genres Mastophora, Celaenia (fig.36,37), Poecilopachys et
justifiant, plus que toute autre,
leur appartenance à une même série
évolutive. Suggéré initialement par les
Robinson (1975)
à
propos de Pasilobus et
repris ensuite par
Eberhard (1980)
et
l’un des précédents auteurs (Robinson,
1982),
ce groupement monophylétique dériverait d’
Orbitèles typiques, incluerait
d’abord Poecilopachys et
l’Araignée
néo-guinéenne Pasilobus,
réunirait ensuite les
Mastophoreae à
bolas (Mastophora, Dicrostichus, Cladomelea)
pour s’étendre enfin aux Celaenieae
qui n'en
produisent pas (Celaenia, Taczanowskia). Aujourd'hui
(2022), suite à divers
travaux plus récents soulignant une même viscosité
de la glue (Cartan & Miyashita, 2000) tous
ces genres sont rattachés à la tribu
des Mastophorini, partie de
la curieuse sous-famille
des Cyrtarachninae.
Le tissu endocrinoïde, à
rechercher, entre
autres intermédiaires, chez Cyrtarachne et
Pasilobus qui en possèdent
vraisemblablement, pourrait bien être, selon A.Lopez, un
caractère anatomique
commun primordial mis en place par la coévolution (évolution convergente...).
Il permet la
maîtrise de proies originales en les attirant avec un ou des
sémiochimiques particuliers, les allomones qui miment les
phéromones sexuelles de leurs femelles. On sait que par
définition, une allomone (du grec ancien ἄλλος allos "autre" et
phéromone), est, selon Brown et Eisner (Brown, 1968) une
substance élaborée et émise par un individu d'une
espèce donnée affectant le comportement d'un membre d'une
autre espèce toute différente au profit du producteur
mais pas du récepteur. La sécrétion du tissu
endocrinoïde stimulerait l'épiderme de Mastophora
libérant dès lors l'allomone.
Il
est à souligner
enfin que malgré une même étrangeté
confinant au
"fantastique", Mastophora reste
complètement isolée, au sein
des Araneidae,
d'un
autre genre extraordinaire, Kaira (Araneinae), plus surprenante
encore
et ne peut à priori, descendre
d’un même ancêtre.
Bien que des proies leur
soient communes (Lépidoptères,
Hétérocères, Diptères) et pareillement
attirées
par des odeurs allomonales
(«mimétisme chimique
agressif»), trop de
caractères
anatomiques et comportementaux s’opposent à un rapprochement des
Mastophorinae
et de Kaira.
Comme le rappelle Levi (1993), ce
dernier taxon est
caractérisé chez le mâle par une apophyse bulbaire
terminale que ne possèdent
pas les précédents ; les cocons
ovigères ont superposés,
dépourvus de couche externe coriace et tissés en
utilisant différemment
les pattes IV ; il
n’existe pas de régurgitations
malodorantes ; les
pattes I et II ne sont pas
fléchies au repos. De plus, les glandes gnathocoxales
(maxillaires)
(Fig.42) présentent un dimorphisme
sexuel,
inexistant chez Mastophora où
il va
de pair avec l’absence de coiffe
embolique dans le bulbe
copulateur.
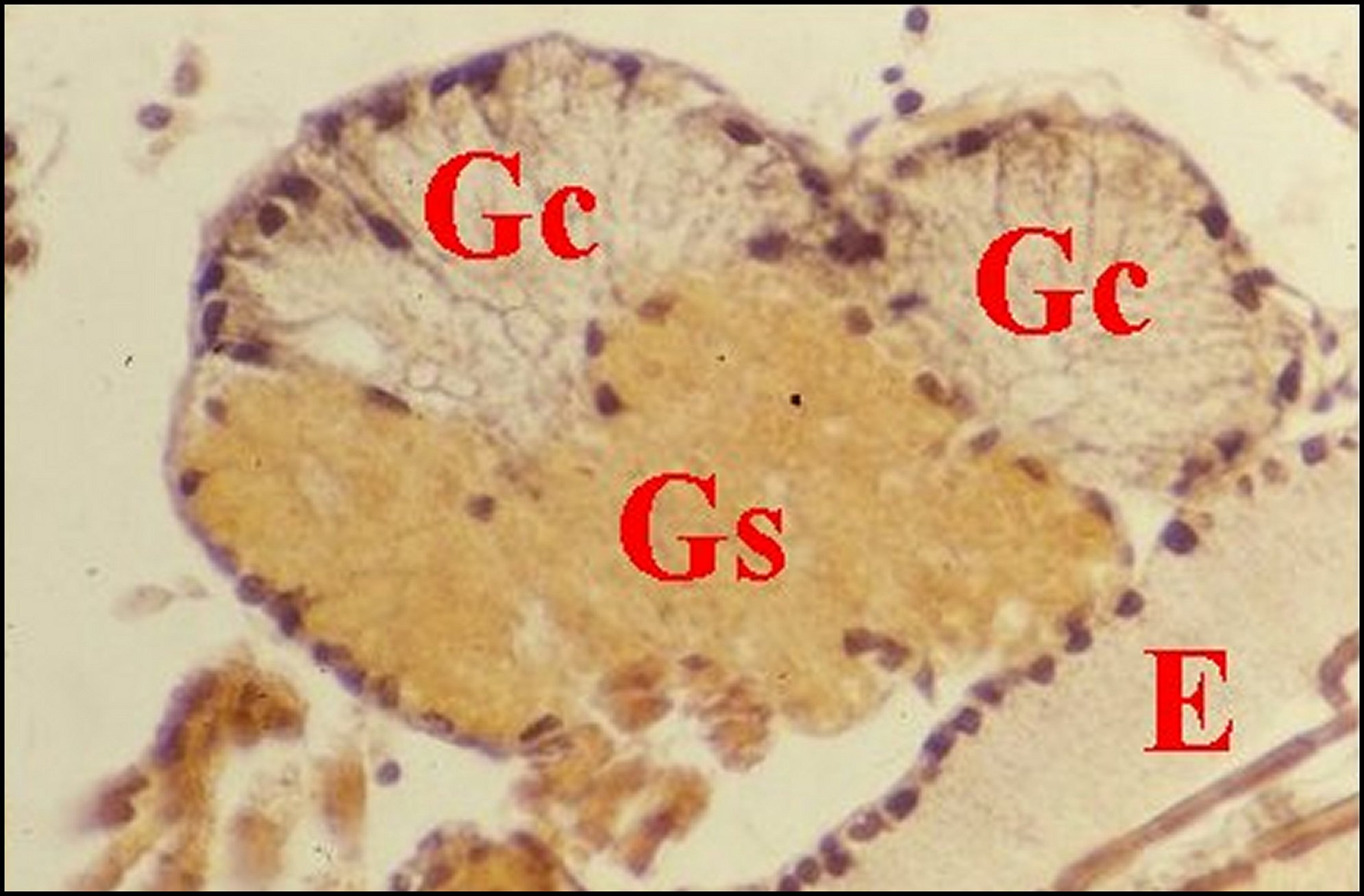 |
| Fig. 39 - Kaira alba mâle : coupe histologique de gnathocoxa. Les glandes "salivaires" sont les unes "classiques" (Gc) et les autres "sexuelles"(Gs), n'existant pas chez la femelle. (A.Lopez,C.H.).Comparer avec la Fig.7: Mastophora mâle. |
La
toile est
rudimentaire mais d’un type et
d’une origine absolument
uniques chez les
Araignées : les
zigzags déposés sur sa
charpente en trapèze des glandessont le
produit flagelliformes et surtout, par leur enduit, de
glandes
agrégées extraordinairement modifiées : les
glandes botryoïdes de Lopez (1986,1998,2001)
 |
| Fig. 40 - Coupe histologique de glande botryoïde (vue trés partielle). |
|
E, épithélium
à petits
adénocytes -Gc, coussinet à cellules géantes
(giganto-cellulaire) - S, glande à soie ampullacée .
Flèches : lumière
(© A.Lopez C.H.)
|
Les proies sont attirées chez Kaira par
ce même enduit «odorant», donc par la toile et les
glandes agrégées
alors
qu’elles sont orientées vers le corps même de
l’Araignée par son tissu
endocrinoïde et l’activation tégumentaire dans le
cas des
Mastophorini.
Pour
conclure, il est regrettable que
malgré son intérêt considérable sur les
plans fonctionnel, phylogénique et
surtout évolutif (coévolution ), le tissu endocrinoïde
abdominal, principal
apport
publié par A.Lopez sur l'anatomie
interne de Mastophora,
ait
été "ignoré" par la plupart des
zoologistes, quand ils ne l'ont pas
délibérément supprimé avec tout son contexte
(in
version française de Wikipédia). Pourtant
les premiers concernés puisqu'il
s'agit de leur propre faune, les
arachnologistes
Nord-américains semblent
essentiellement orientés vers
la chimie des composés
volatiles, non leur origine précise (Yeargan
& Quate,1996,1997; Stowe,1986 ; Hutchinson,1903).
| "ARAIGNEES" |