|
Tissu
endocrinoïde des Mastophorini et autres Cyrtarachninae
ANATOMIE
DES
ARAIGNEES :
VINGT-CINQ ANS
DE RECHERCHES
(Version 2022) |
En
rapport
étroit avec Mastophora
sauvegardé Cyrtarachninae
Araignées
et allomones
et Kaira
| Les Cyrtarachninae et notamment celles du
genre Mastophora sont des Araignées extraordinaires, de la grande
famille des Araneidae, qui attirent leurs proies, le plus souvent des
Papillons Hétérocères mâles, en émettant des allomones, substances
attractives
simulant les phéromones sexuelles femelles de ces Lépidoptères
(Mimétisme chimique agressif). Selon André Lopez qui l'a découvert par
histologie, du moins dans trois genres bien distincts, un étrange
organe abdominal baptisé "tissu endocrinoïde" serait à l'origine du
processus, une nouvelle preuve de l'évolution darwinienne. Ce
tissu est une structure anatomique étonnante située entre les glandes à
soie agrégées, constituée par des amas cellulaires en ilôts,
cordons
ou rubans, beaucoup plus développés chez la femelle et montrant
parfois une fausse lumière centrale. Les cellules, anguleuses, ont des
noyaux arrondis et des cytoplasmes acidophiles
montrant des signes ultrastructuraux de production
stéroïdienne. Il est probable qu'elles
stimulent le tégument
abdominal comme les glandes de mue
et les ecdystéroïdes induisant ainsi la production d' allomones
attirant les mâles d' Hétérocères dans le cadre du mimétisme chimique
agressif. |
|
Couleurs
conventionnelles :
En noir et italiques, termes anatomiques ; en violet,,,noms
génériques et spécifiques ; en vert,,noms de
familles et sous-familles; en
orange,, parties
les plus importantes
et résumés ; en bleu, liens divers.
Abréviations
conventionnelles :
M.E.B.
:
(photographie en) microscopie électronique à balayage
C.H. : coupe histologique (microscopie photonique) |
1 - Introduction
1-1 - Morphologie
1-2- Répartition
1-3- Systématique,
Phylogénie
2 - Anatomie interne
2-1-Abdomen
2-1-1 -
Tissu abdominal endocrinoïde
2-1-1-a
- Histologie
3 - Commentaires
3-1 - Considérations anatomiques
3-2 - Considérations physiologiques
3-3 - Considérations phylogéniques
Au
point de vue morphologique,
toutes les Mastophorini
(Araignées Araneidae Cyrtarachninae) se
singularisent
par des pattes épineuses et
un corps trapu.
Leur abdomen
est globuleux, avec des bosses
dorso-latérales parfois
très saillantes et des teintes cryptiques (Fig.4,5) ; le céphalothorax
montre de
curieuses protubérances
dorsales : en forme
de «couronne», de « mitre » et, dans le cas des Mastophora ,
soit d’une paire
de «cornes occipitales» plus ou moins
bifurquées (Fig.1 à 3 : flèches et
"H"),
soit de files de pointes
acérées (Fig.1).
 |
 |
 |
|
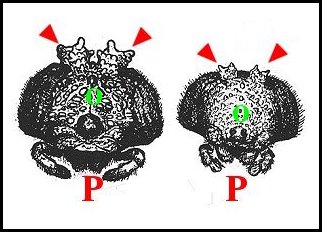
Fig.1 - Mastophora
cornigera et bisaccata ,
femelles. Prosomas vus de face (d'après Gertsch)
|
Fig. 2 -M.
cornigera, mâle mûr : vue dorsale totale
|
Fig.3 -Vue dorsale du prosoma sans pattes ni palpes |
|
H et
flèches rouges, cornes bifurquées
; O, yeux ; P, pédipalpes. Fig.3 : © A.Lopez M.E.B. |
||
Les autres Cyrtarachnineae ont aussi un
corps très
ramassé, avec l’abdomen
globuleux souvent garni de tubercules ou
protubérances
épineuses et paré de
couleurs vives (Poecilopachys) (Fig.4 à 6
) ou déprimé transversalement (Pasilobus) (Fig.7).
 |
 |
 |
 |
|
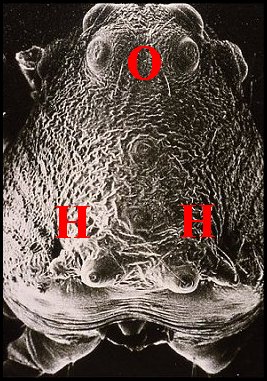
Fig.4 - Mastophora
cornigera
Femelle, vue dorsale. Flèche
: cornes
|
Fig. 5 - Celaenia excavata Femelle, vue dorsale (Col.A.Lopez) |
Fig.6-
Poecilopachys australasia Femelle, vue dorsale (Col.A.Lopez) |
Fig.7-
Pasilobus sp. Femelle,vue frontale (Col.A.Lopez) |
1-2 - Répartition
Au point de vue biogéographique, les Mastophorini ont une répartition tropicale et sub-tropicale très étendue : Amérique, du Sud des USA jusqu’à l’Argentine ; Afrique australe ; région australienne (Australie, Nouvelle-Zélande). Les Cyrtarachneae sont une tribu d’Araignées tropicales propres à l’Ancien Monde (Australie, Nouvelle-Guinée, Inde, Chine, Japon).
1-3 - Systématique, Phylogénie
Au
point de
vue systématique, les Mastophorini
comportent deux tribus : les Mastophoreae, avec
les genres
américains Mastophora
et Agatostichus, africains Cladomelea
et Acantharachne,
australo-néozélandais Dicrostichus et
Ordgarius ;
les Celaenieae, avec
seulement deux genres,
Taczanowskia, sud-américain,
et Celaenia
de la région
australienne.
La
tribu des Cyrtarachneae (Simon,1892)
réunit
quelques genres peu connus encore: Cyrtarachne, hindou et
extrême-oriental ;
Poecilopachys et Pasilobus, austro-hindous
; Paraplectana, extrême-oriental.
Au
point de vue phylogénique, les Mastophorinae
forment un groupe assez
homogène,
anciennement
détaché des Araneidae dont
il
possède toutefois les
caractères
anatomiques fondamentaux. De même, les Cyrtarachneae
dérivent manifestement de la même
famille, par
leurs morphologie et industrie
séricigène.
1-4 - Comportement de chasse
Au
point de vue éthologique, les Mastophoreae
élaborent de fils pendants
garnis d’un (Fig.9,10) ou plusieurs (Fig.8)
globules
visqueux que
les femelles font tourner avec l'une des
pattes ambulatoires en un mouvement
de manège ou tourniquet («whirligig»)
(Fig.9), les
manipulent
ainsi comme des «bolas» de gauchos
sud-américains («bolas-spiders»),
engluent
les proies à la volée et halent ensuite le fil qui en est
chargé(Fig.11)
pour
les
consommer après paralysie. Les "bolas»
inutilisés
sont renouvelés
périodiquement par ingestion.
L’ordre de la patte varie d’ailleurs
selon
le genre : I chez
Mastophora et Agatostichus, II chez Dicrostichus
et Ordgarius,
III chez Cladomelea.
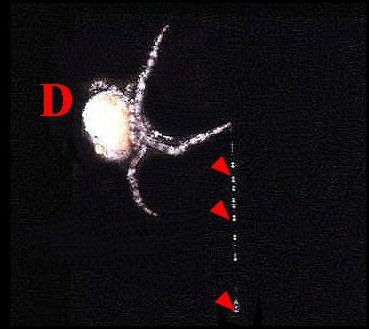 |
 |
| Fig.8 - Dicrostichus magnificus femelle (D) tenant un bola à plusieurs globules (flèches) (Col.A.Lopez) | Fig.9 - Mastophora faisant tourner son bola à globule unique : séquences du mouvement par artifice photographique. |
 |
 |
| Fig.10 - Mastophora bisaccata femelle (M) tenant un bola à globule unique (flèche) et attirant un Hétérocère (L). | Fig.11- La même vient d'engluer l' Hétérocère avec le globule de son bola sous tension (flèche) |
Les Celaenieae
(Fig.5),
en revanche, ne produisent aucun
appareil soyeux de
capture et saisiraient
directement les proies avec
leurs pattes I et
II étendues
(Mc Keown,1952 ; Eberhard,1981).
Pour
leur part, les Cyrtarachneae
tissent
des toiles
orbiculaires modifiées, asymétriques,
larges,
triangulaires ou polygonales, plus ou moins inclinées sur
l’horizontale, pourvues d’un petit nombre de radii et ne
comportant pas la classique
spirale gluante des Araneideae (Fig.12,13). Cette
dernière est remplacée par un «fil visqueux»
(«viscid line»)
lui-même formé de «fils en pont»
(«spanning
threads») lâchement tendus dans les secteurs (Clyne,
1973 : Poecilopachys)
(Fig.12). Leur partie
centrale
est garnie d’un matériel très gluant, en larges gouttelettes et l’une
de leurs extrémités se rompt facilement lorsqu'un Insecte
(Lépidoptère) y adhère,
pendant
alors comme un bola
chez Pasilobus
(Robinson & Robinson, 1975)
(Fig.13).
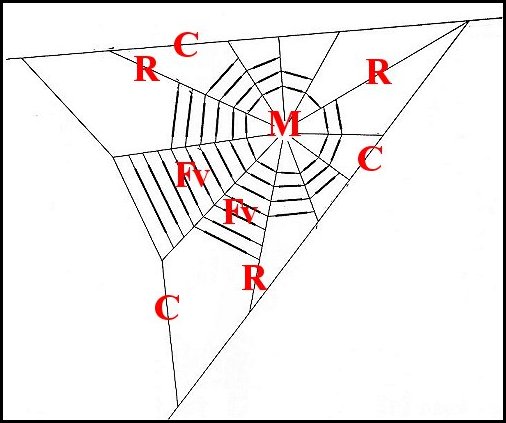 |
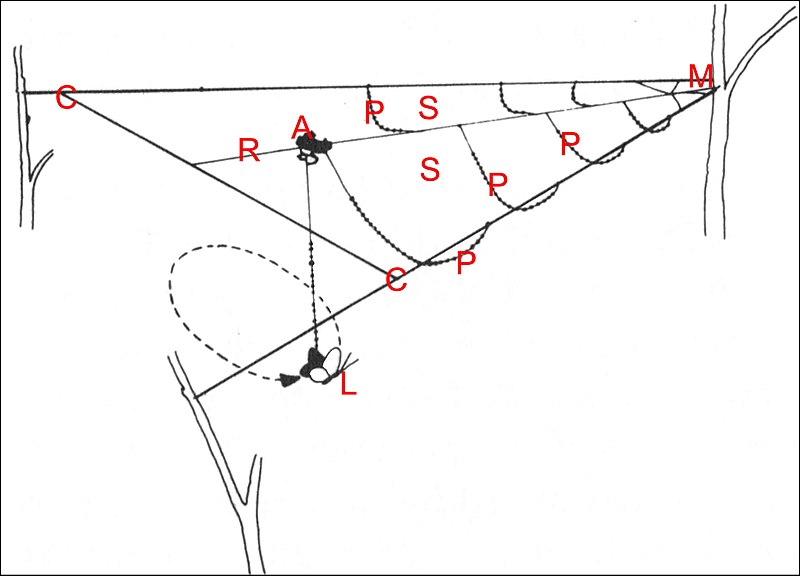 |
| Fig.12.-
Toile
de Poecilopachys
(d'après Clyne) |
Fig.13- Toile de Pasilobus (d'après Robinson & Robinson) |
| C, charpente ; Fv, fil visqueux ; M, moyeu ; R, radius | P, Araignée ; L, Lépidoptère. Flèche :"fil en pont"rompu et pendant. |
Une autre singularité biologique fort remarquable est décrite dans les genres Dicrostichus (Lowry, in Mc Keown,1952) et Mastophora (Gertsch,1947) : la maturité sexuelle précoce des mâles nains . Ces derniers sortent adultes du cocon ovigère, leurs bulbes copulateurs bien développés permettant de les identifier aisément dès l’émergence et de ne pas les confondre avec de petites araignées venant d'éclore ("spiderlings").
Sa présentation sera réduite à celle de l'abdomen, le prosoma
de Mastophora n'étant étudié que dans cette partie du site
(glandes à venin, gnathocoxales, diverticules intestinaux dans les
"cornes",
L'anatomie
interne de la sous-famille entière
étant inconnue jusqu'à ses travaux, A.Lopez
(1985a) a donc recherché dès
cette date si des
particularités histologiques peuvent expliquer celles de la morphologie
externe, du comportement prédateur et du développement
post-embryonnaire. il a utilisé comme
matériel d'étude les quatre Araneidae
Mastophora
cornigera Hentz et M. bisaccata
Emerton (Fig.1à4, fig.10,11) (Mastophoreae), Celaenia excavata
(Celaenieae) (Fig.5) et
Poecilopachys
australasia (Cyrtarachneae)(Fig.6)
Les deux
premiers taxons étaient
représentés
par des exemplaires à divers stades évolutifs, y compris
des jeunes femelles et tous les mâles mûrs sortis depuis peu du cocon (M.bisaccata) ;
ils provenaient des USA, respectivement de Californie (Fiesta Island,
San Diego) et de Floride (Gainesville, Alachua) où
M.K.Stowe les a
récoltés et fixés (1984, 1991-92). A.Lopez les a ensuite préparés pour
l’histologie selon
des
techniques banales ; le matériel destiné à la M.E.T à été
inclus
en Floride et examiné ultérieurement dans le Laboratoire ariégeois
de Moulis (Note1).
Les
exemplaires de Celaenia sp. se réduisaient à
deux femelles adultes provant de Nouvelle-Zélande (Takapuna, Auckland)
où G.Lowe les a capturées et fixées en 1991.
Poecilopachys australasia, une femelle adulte, a été récoltée et fixée en Australie (Brisbane, Queensland) par Benoit Lopez, fils de l'auteur (1994).
Ce dernier a préparé la majeure partie du matériel pour l’histologie, selon des techniques banales ; une Mastophora bisaccata, comme par ailleurs Kaira alba, ont seuls fait l’objet d’un examen au M.E.T. dans le Laboratoire ariégeois (Note1).
La documentation sur Celaenia et surtout Poecilopachys
ne
concerne que certaines glandes
à soie, le tissu
endocrinoide et la partie ventrale du céphalothorax.
Il renferme notamment l'appareil génital présenté chez Mastophora et surtout l' appareil séricigène également décrit en détails dans le même genre
▬ Les glandes ampullacées, au nombre de 4 se terminant dans les filières antérieures et moyennes.
▬Les glandes piriformes,
«mi-parties» selon l’expression de Millot aboutissant
aux filières antérieures.
▬Les glandes
agrégées, de volume
considérable (Fig.14),
multilobées,
elles ont une
lumière anfractueuse, un aspect
arborescent et des canaux
excréteurs, noduleux et
à plusieurs couches
pariétales aboutissent
aux filières
postérieures.
▬Les glandes
flagelliformes rejoignent filières
postérieures par leurs canaux
excréteurs où chacun d’eux s’associe
étroitement en « triade fonctionnelle » avec
une paire de canaux de glandes agrégées.
▬Les
glandes
tubuliformes se terminent dans les filières
moyennes et postérieures.
L’appareil
séricigène du mâle
est réduit à 4
catégories : glandes
ampullacées majeures et
mineures, glandes
piriformes, glandes
aciniformes.
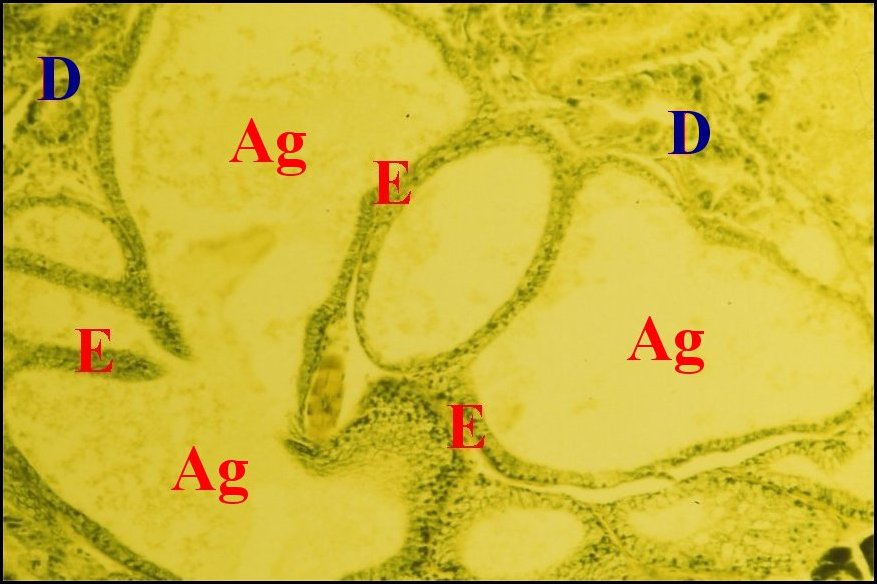 |
| Fig.14 - Mastophora cornigera, femelle adulte. Vue très partielle de l'abdomen montrant le grand développement des glandes à soie agrégées. |
| Ag, agrégées - D, diverticule chylentérique - E, épithélium des agrégées (© A.Lopez C.H ) |
De son
côté, Caelenia
excavata parait
totalement
dépourvue de glandes
agrégées,
catégorie
dont l'absence est connue, par ailleurs chez d'autres Araneidae construisant
pourtant
une
grande toile (Cyrtophora : Kovoor & Lopez,1982)
Par ailleurs, chez Poecilopachys
australasia,
les mêmes glandes ont un
développement considérable. Leurs canaux
présentent une
paroi à nodules si
étendus qu'ils semblent
parfois s'en
être isolés pour former un tissu distinct
(Fig.15).
 |
| Fig. 15 - Poecilopachys australasia, femelle : partie de l'appareil séricigène |
| Glandes séricigènes agrégées et piriformes. D, canal excréteur de glande agrégée - N, nodules externes - Pi, glandes piriformes. Flèches : cuticule (A.Lopez,C.H.). (© A.Lopez C.H.). |
2-1-1 -Tissu abdominal endocrinoïde
Au
voisinage
immédiat des agrégées
et ampullacées
des
Mastophora, de Celaenia, et des agrégées de Poecilopachys
australasia,
il existe un tissu très
particulier , d’aspect glandulaire mais sans canaux excréteurs,
entouré d’hémolymphe, ne
contractant que des
rapports de contiguïté avec l’appareil
séricigène et les diverticules
chylentériques.
Dans
le cas de Mastophora
tout d'abord, il
est réduit chez
le mâle (Fig.16,17), bien plus
développé chez les femelles immatures du
même genre (Fig.18,20,21)
et atteint
un volume considérable
lorsqu’elles deviennent adultes (Fig.19).
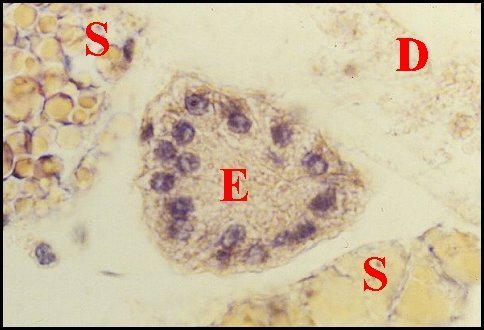 |
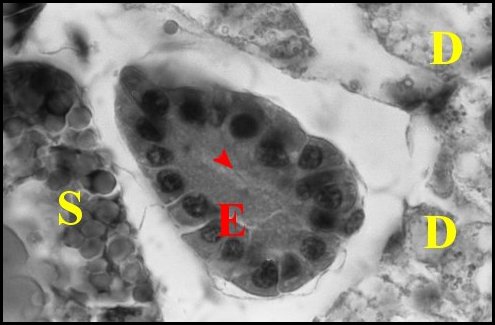 |
| Fig.16 - Mastophora cornigera, mâle : ilôt de tissu folliculaire endocrinoïde. | Fig.17 - Même mâle :
autre ilôt de tissu
folliculaire endocrinoïde(photo noir et blanc). |
| E, tissu - D, diverticule
intestinal - S, glande à soie. Flèche :
cavité
centrale (© A.Lopez
C.H.) |
|
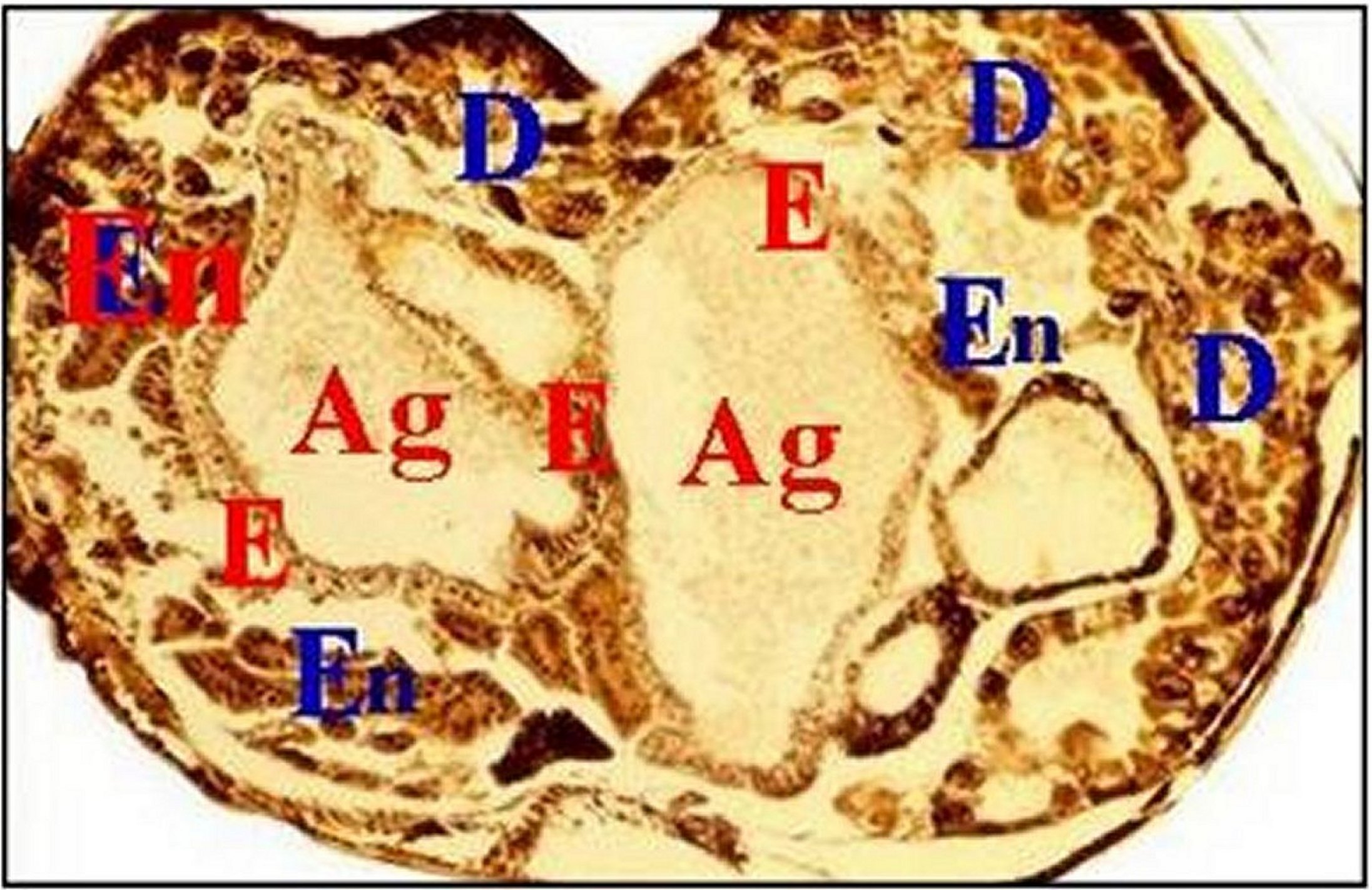 |
|
Fig.18
- Mastophora
cornigera, femelle immature. Coupe
transversale et totale d'abdomen
|
| Ag, agrégées - C, canal excréteur d'agrégée - D, diverticule chylentérique - E, épithélium des agrégées - En, tissu endocrinoïde (© A.Lopez C.H ) |
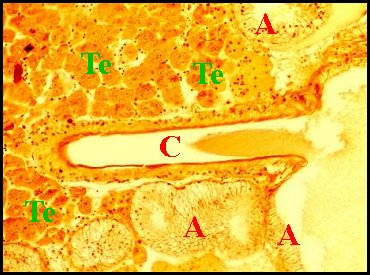 |
| Fig.19 - Mastophora cornigera, femelle adulte : vue trés partielle d'une coupe d'abdomen. |
| A,
corps de glandes agrégées - C, leur canal excréteur - Te, tissu
endocrinoïde
folliculaire (© A.Lopez C.H.) |
Dans tous les cas, ses cellules
sont
bien individualisées, de tailles et de
forme assez constante globuleuse, polyédrique, cuboïdale ou même
conique. Leur cytoplasme
est
acidophile, finement grenu, parfois criblé de très petites vacuoles.
Leur noyau
est arrondi, clair, nucléolé, riche en chromatine
bien visible.
Elles
se réunissent
en petits massifs et cordons peu sinueux, arrondis dans les coupes
transversales, compacts ou montrant souvent une cavité axiale très
étroite, presque
virtuelle et qui n’est pas une
vraie lumière
(Fig.20,22,23).
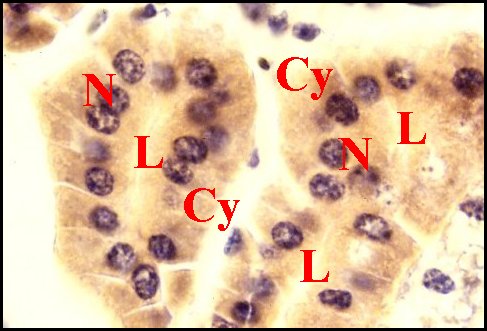 |
 |
| Fig.20 - Mastophora cornigera, femelle immature : détails du tissu endocrinoïde folliculaire. | Fig.21
- Autres détails du tissu endocrinoïde folliculaire
dans une partie différente de l'abdomen. |
| Cy, cytoplasmes de cellules endocrinoïdes - I, cellules à ferment de diverticule intestinal - L, cavité ("lumière") centrale - N, noyaux de cellules endocrinoïdes (© A.Lopez C.H.). | |
Ces groupements
de cellules sont
assez peu nombreux,
bien
isolés les uns des autres de taille
relativement
grande chez les immatures (Fig.20,21),
en quantité beaucoup plus grande, tassés les uns contre les autres
et plus petits chez les femelles adultes (Fig.19,22,23).
Ils se présentent comme des structures sacciformes collabées,
entièrement closes, dépourvues de toute structure
canalaire, baignées directement
par l’hémolymphe.
Un tel ensemble de caractères est très évocateur de leur nature
«endocrinienne» et n’est pas sans évoquer les follicules
(vésicules)
thyroidiens de Vertébrés.
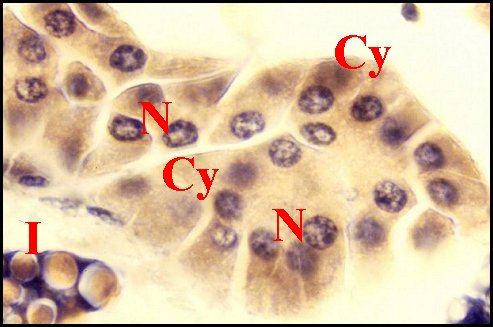
 |
 |
| Fig.22- Mastophora cornigera, femelleadulte : détails du tissu endocrinoïde folliculaire. |
Fig.23
- Autres détails du tissu endocrinoïde folliculaire
dans une partie voisine de l'abdomen.
|
| N, noyaux des cellules endocrinoïdes disposées en ilôts juxtaposés - V, vaisseaux. Les flèches désignent des cavités ("lumières") centrales (© A.Lopez C.H.). | |
Faute d’une
meilleure appellation, le tissu
peut
donc être qualifié de «folliculaire
endocrinoïde». Il
s’insinue entre les organes
voisins,
notamment les lobes des
agrégées et les diverticules
chylentériques (Fig.18,19)
qui semblent
d’ailleurs les déformer par compression. De plus, il tend à se répartir
en
quatre amas : deux
ventraux,
paramédians, situés un peu en arrière du niveau des phyllotrachées (poumons), et
deux latéro-dorsaux,
très externes. Les amas sont seulement accolés aux glandes séricigènes
agrégées, entourent
l’origine de leurs canaux
excréteurs (Fig.19).
Le
tissu
folliculaire endocrinoïde du
mâle de(Mastophora
cornigera
se limite à deux petits massifs
cellulaires
symétriques pouvant montrer
quelques signes de dégénérescence :
condensation ou vacuolisation
exagérées du cytoplasme,
pycnose nucléaire (Fig. 16,17).
Le tissu endocrinoïde
se retrouve chez
la
femelle de Celaenia
excavata
où ses ilôts se logent entre les glandes
tubuliformes, qui tendent à
les
comprimer (Fig.24,25).
Ils montrent, eux aussi, des cavités centrales trés étroites que peut
limiter une seule assise
cellulaire (Fig.24).
 |
 |
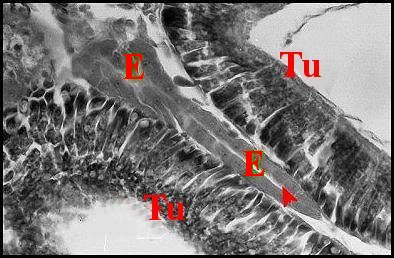
| Fig.24-
Celaenia excavata, femelle
:
vue trés partielle d'une coupe d'abdomen (photo noir et blanc). |
Fig.25 - Autre ilôt endocrinoïde dans une partie différente de l'abdomen, détail (photo noir et blanc). |
| Tu, glandes
tubuliformes - E, ilôts de tissu endocrinoïde
folliculaire. La flèche désigne une cavité centrale en fente. (© A.Lopez C.H.) |
|
Chez
la femelle de Poecilopachys
australasia, il
présente aussi
un développement considérable. Insinué entre les lobes
des
énormes glandes agrégées, il se compose
encore de massifs cellulaires en ilôts éosinophiles montrant de gros noyaux ronds vésiculeux, nettement
nucléolés, et de
rares ébauches de cavités centrales (Fig.26,27).
 |
 |
|
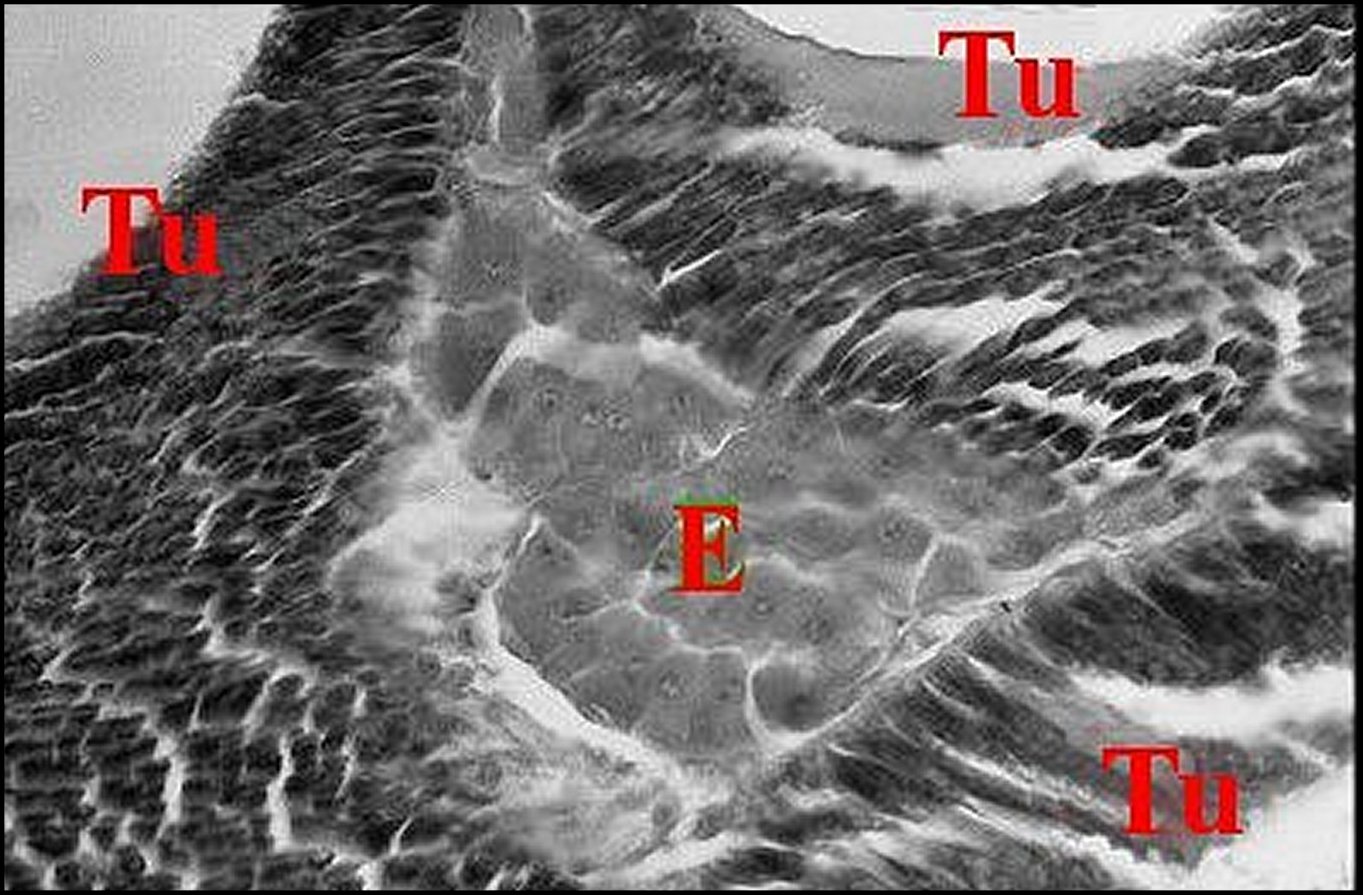
Fig.26
- Poecilopachys
australasia femelle : coupe
d'abdomen. .
|
Fig.27 -Poecilopachys australasia, même femelle : autre coupe d'abdomen.
|
| Ag, glande agrégée (corps) - E, tissu endocrinoïde (© A.Lopez C.H.) | |
2-1-1- b Ultrastructure
Elle n'a été
étudiée (sommairement) que chez Mastophora
bisaccata.
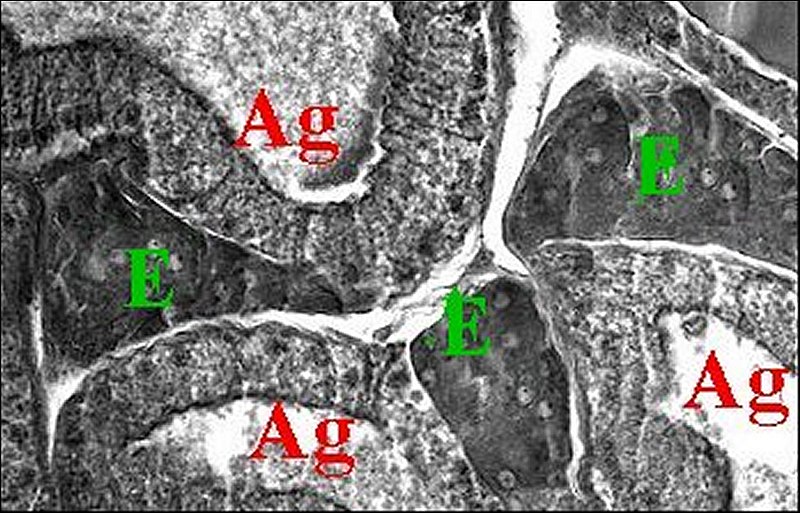
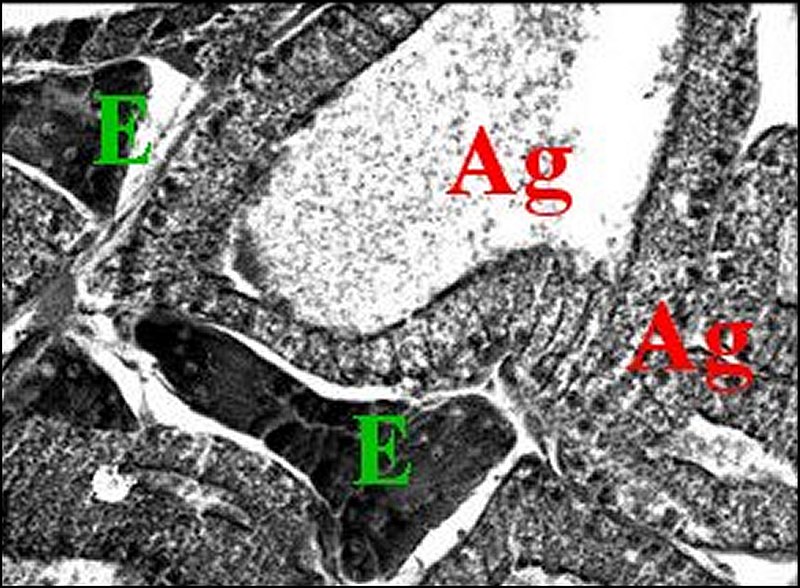
Les amas cellulaires sont entourés par une lame basale mince formée d’un matériel finement granuleux (Fig.28).
Il semble exister une discrète polarité cellulaire, notamment dans les amas « évidés » en leur centre car le plasmalemme tend à former des invaginations intracytoplasmiques au-dessous de la basale.
Le noyau, clair et arrondi, renferme de la chromatine dispersée dans son nucléoplasme et formant aussi de grosses mottes marginales.
Le réticulum endoplasmique est remarquablement développé, lisse, et se compose de cisternae pouvant se dilater en chapelets vésiculaires.
Les
autres organites sub-cellulaires sont des mitochondries allongées
pouvant présenter
des crêtes tubulaires,
de petits dictyosomes,
des lysosomes, des
polysomes et
des microtubules épars
(Fig.29).
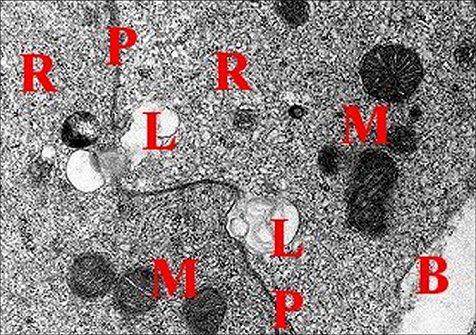 |
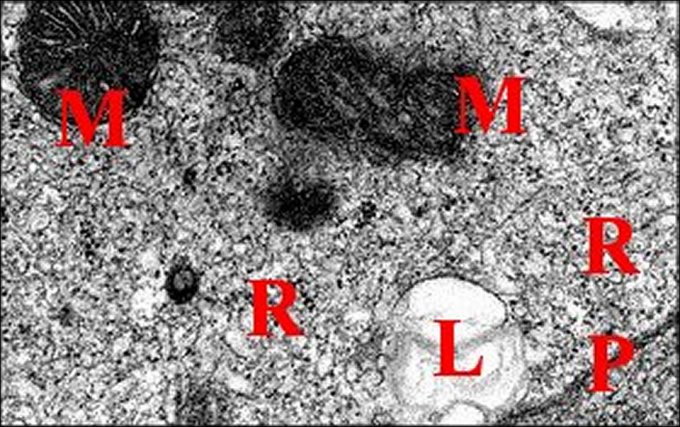 |
| Fig. 28 - Mastophora bisaccata : cellules du tissu endocrinoïde folliculaire. | Fig.29 - Mastophora, la même, tissu endocrinoïde : détails des organites |
| B, lame basale - L, lysosomes - M, mitochondries à crêtes tubulaires- P, plasmalemme - R, réticulum lisse (© A.Lopez M.E.T.) | |
L' appareil
séricigène de Mastophora (immatures, femelles adultes,
mâles mûrs), peut être
aussi celui de Poecilopachys
(non étudié
en
détails), a une
composition et
une structure d’ensemble qui sont typiques des Araneidae,
et ne
refléte donc pas
la disparition de la toile orbiculaire classique. Les glandes
flagelliformes produisent
vraisemblablement
les fils axiaux pelotonnés des "bolas" de Mastophora
(fig.2) et la
fausse spirale de Poecilopachys (Fig.12)Clyne,1973) Dans les
deux genres, le grand
développement des agrégées assure une sécrétion de «glue» très abondante, ce qui
garantit la production répétée des globules de «bolas» et
la mise en
place des enduits visqueux centraux sur les « fils en pont » de la fausse spirale.
En revanche,
l'appareil séricigène de Celaenia
semble bien être dépourvu de glandes agrégées, ce qui
explique un
comportement de chasse excluant tout fil de capture.
Dans aucun de ces cas, les agrégées,
lorsqu’elles
existent, et
toutes les autres catégories
glandulaires
ne
comportent aucune cellule spéciale qui puisse
être
comparée à celles des glandes botryoïdes chez
Kaira
alba.
De plus, il n'existe aucun
rapport de continuité entre les glandes à soie, notamment les agrégées ou
arborescentes, productrices de la glue, et le tissu folliculaire
endocrinoïde abdominal
Ce dernier ne peut être
confondu
avec aucune autre structure histologique, notamment la glande de mue.
Il est en effet bien
différent du tissu
interstitiel adipoïde
étroitement associé avec l’intestin
(Millot, 1926),
ne
peut
correspondre à des glandes
séricigènes ébauchées ou
involuées et paraît aujourd’hui ne plus pouvoir être considéré comme un
simple
contingent
postérieur de la glande de mue (Fig.30), première conception
erronée de Lopez
(1985a) chez Mastophora
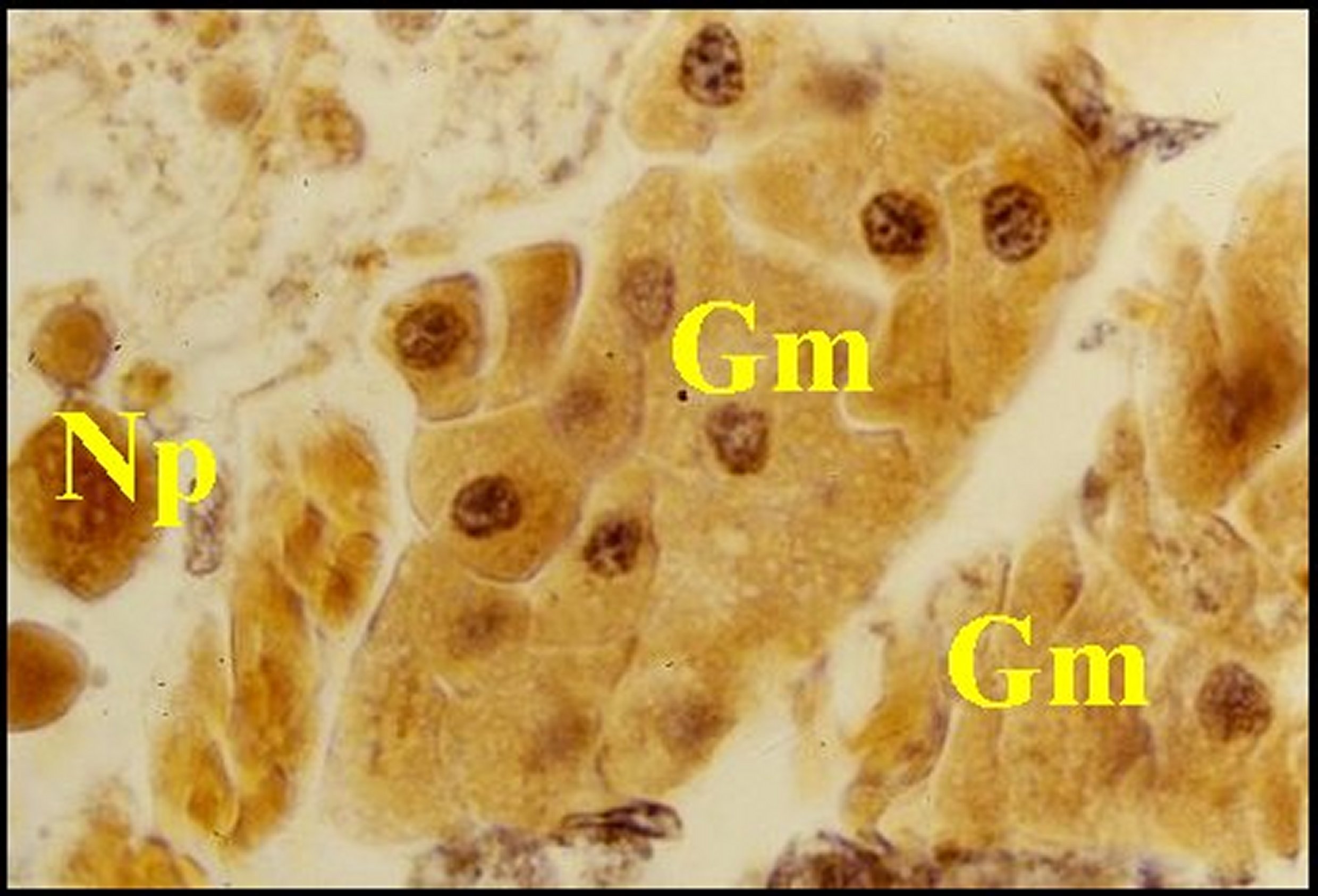 |
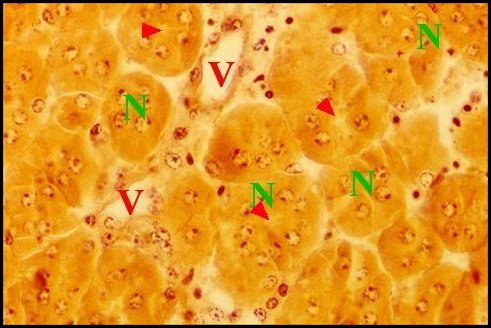
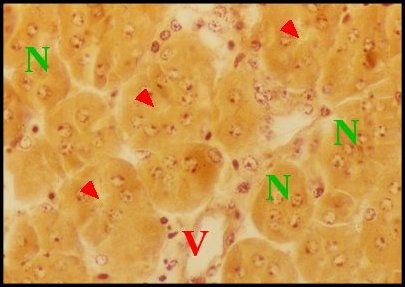
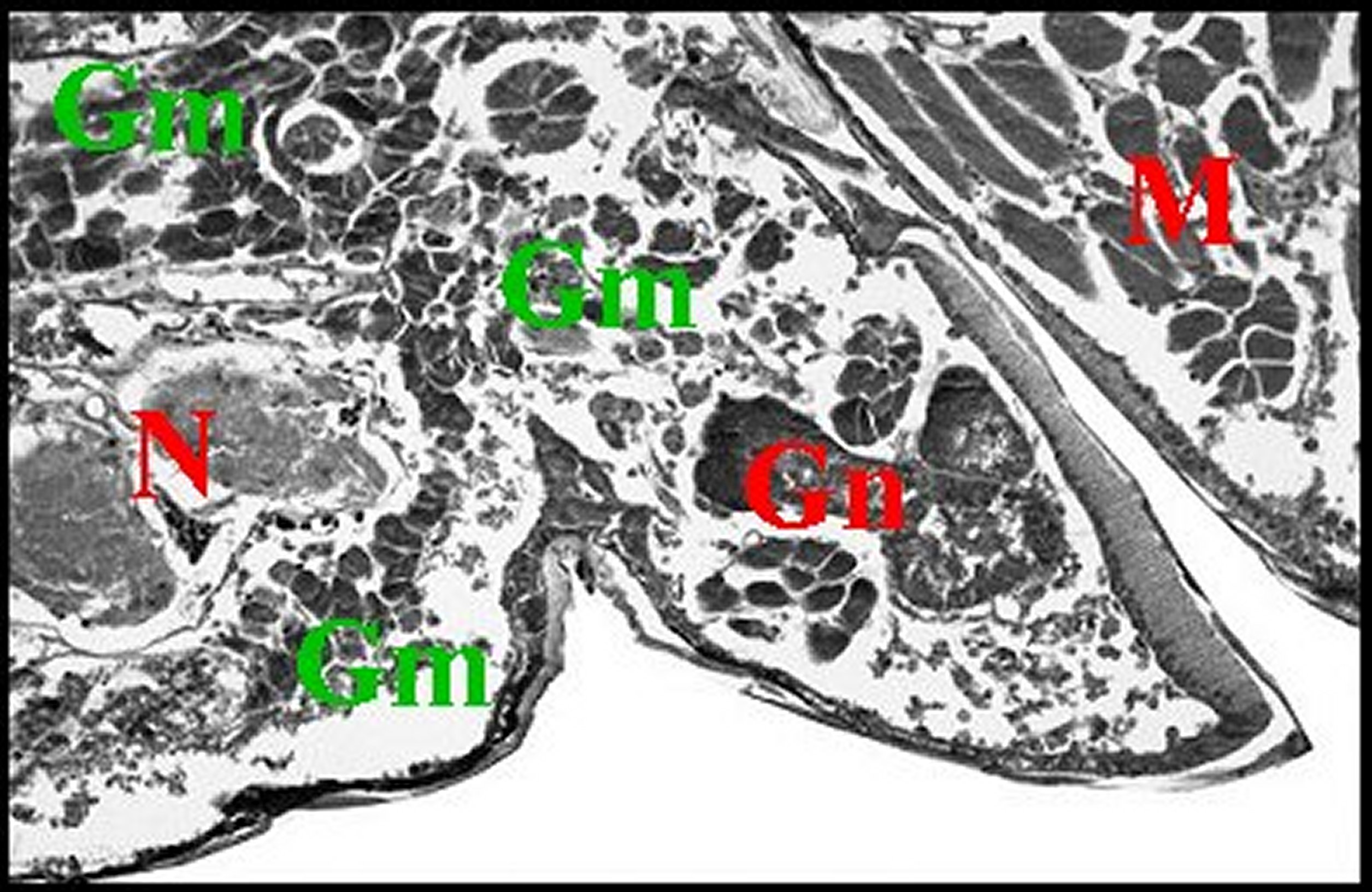 |
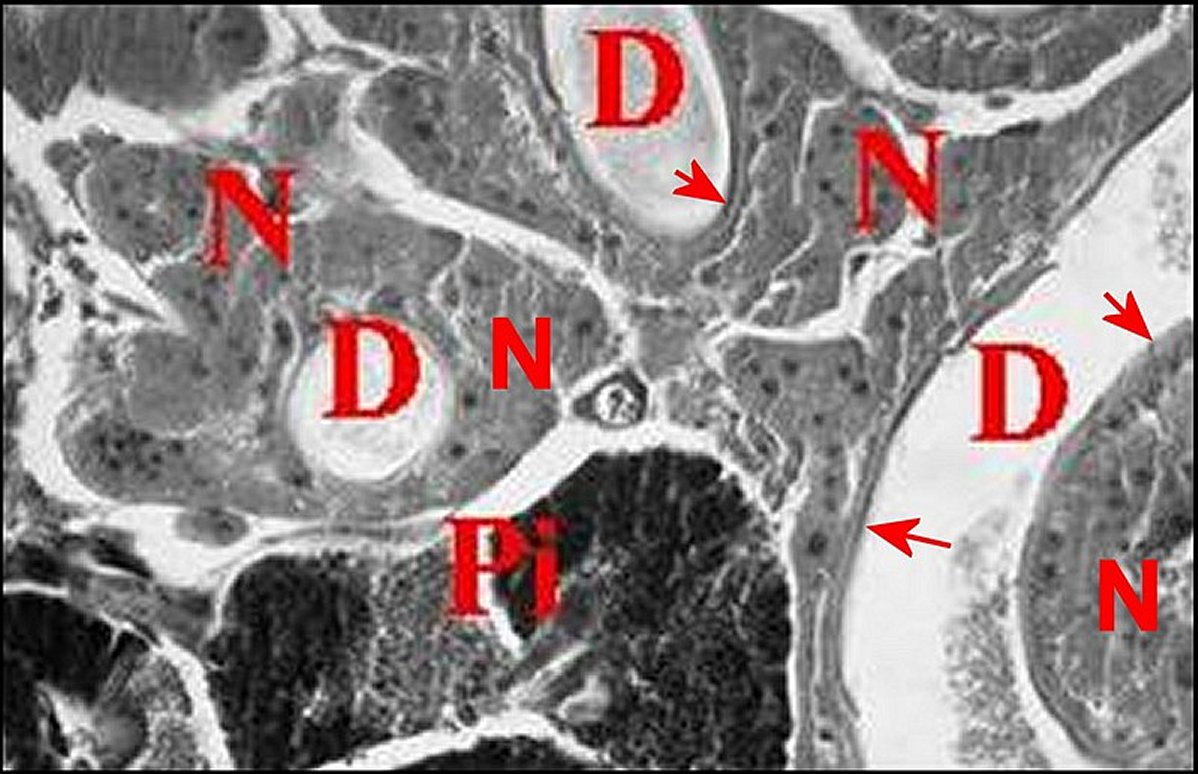
| Fig. 30 - Mastophora cornigera, glande de mue (Céphalothorax). | Fig.31 - Poecilopachys australasia, glande de mue (Céphalothorax). |
| Gm, glande de mue prosomatique
- Gn, gnathocoxe
avec ses
glandes "salivaires" - M, muscle - N, ganglions nerveux - Np, néphrocyte (© A.Lopez C.H.) |
|
Découverte
et décrite par Millot
(1930) sous le nom de"tissu
endocrine",
cette dernière
présente dans
tout l’ordre des Araignées
une stricte localisation céphalothoracique (Streble,1966 ;
Bonaric,
1980) à l’exception toutefois de certaines Mygales
dont les Liphistiidae
(Lopez,1981) où on la retrouve
jusque dans
l’abdomen au contact des
gros nerfs (Legendre,
1981). Son rôle
dans les
mécanismes de l’ecdysis
a été évoqué par Legendre
(1959) qui la considère comme un homologue mésodermique
de
celle des
Insectes,
et
confirmé par Bonaric
(1980) qui
lui a attribué son nom définitif.
Cette glande de mue
(Fig.30,31)
adopte dans le prosoma une disposition spatiale qui
est en grande partie métamérique («organe antérieur» et «organe
postérieur», «groupes latéraux») alors que le tissu
folliculaire
endocrinoide
abdominal ne
semble pas en présenter une. De plus, les derniers examens
comparatifs
de Lopez montrent que leurs aspects histologiques ne sont pas
exactement
superposables. Contrairement au tissu
folliculaire endocrinoïde de Mastophora,
Celaenia
et Poecilopachys, les cellules
de
la glande de mue ont une
taille assez variable
et une forme plus irrégulière. Elles se groupent plutôt en rubans et
en
travées compactes où il n’y a pas de «lumière» ; leur
association
avec des néphrocytes et
des hémocytes est
quasi-constante (Fig. 30,31).
Les caractères
ultrastructuraux sont en revanche assez voisins (réticulum endoplasmique
lisse développé,
crêtes mitochondriales
tubuleuses).
Sur le plan
fonctionnel, il est hors de doute que le tissu folliculaire
endocrinoïde abdominal ne soit impliqué dans
l’attraction et
la capture de
proies, le
produit des régurgitations
de Mastophora et l'épiderme
de ses pattes étant
à exclure, du moins chez les
femelles adultes.
Ses relations avec la substance volatile attractive autorisent deux hypothèses.
▬ La
plus séduisante est inspirée par le
mode d’action des ecdystéroïdes, dont on sait qu’ils sont
responsables du
déterminisme de la mue et de autres processus mettant en jeu la
différenciation
cellulaire (métamorphoses, formation des œufs) aussi bien chez
les Insectes que
chez les Araignées. Le tissu folliculaire endocrinoide
présentant des indices
ultrastructuraux d’élaboration stéroïdienne, ses
sécrétions pourraient activer
les propres cellules
épidermiques corporelles de Mastophora tout comme Celaenia y
stimulant, non plus la formation d’une cuticule nouvelle comme dans les
mues
mais la synthèse d’ARN, proteines et polysaccharides,
peut-être aussi
d’enzymes, nécessaire elle-même à celle des
attractifs volatiles et "odorants" sans aucun doute des allomones. L’absence
d’images
histologiques concrètes en faveur d’une participation du
tégument dans des
zones spécifiques (cornes , surface de l'abdomen) n’est certes
pas en faveur de
cette théorie. Il est prouvé cependant que chez les
Papillons femelles eux-mêmes,
certains organes abdominaux dits « appelants » (sacculi
laterales et
autres glandes évaginables) ont des cellules
épidermiques
banales que recouvre
une cuticule ordinaire (Steinbrecht,
1964).
Toutefois, les observations d’auteurs nord-américains tels qu’ Eberhard (1977) privilégient le corps même de l’Araignée et excluent formellement une quelconque attraction par les produits des glandes à soie, confirmant ainsi la première hypothèse de Lopez.
L'implication finale d'allomones montre qu'il s 'agit bien là d'un magnifique exemple de mimétisme chimique agressif.
3-3- Considérations phylogéniques
Ainsi
que
Lopez
(1985b) l’a déjà évoqué au début de ses
observations sur les «Araignées excentriques» du nouveau Monde et
confirmé ensuite ultérieurement (Lopez,1998),
il existe une ressemblance certaine
entre les
Mastophora, Celaenia et
Poecilopachys
non
seulement par leur équipement séricigène (composante agrégée
«arborescente» en particulier) mais surtout, par le tissu
folliculaire
endocrinoïde
abdominal dont elles sont pourvues.
Ce dernier a été malheureusement ignoré par Levi
(2003) dans sa mise au
point sur le premier genre
(Note 2).
Quelques
années
plus tôt,
Eberhard (1980) dans son étude sur Mastophora
dizzydeani et
Robinson
(1982), dans sa
monographie sur les Araignées
de Papouasie, avaient proposé de
rattacher les Mastophoreae,
les Celaenieae et les Cyrtarachneae, dont Poecilopachys,
à
un même groupe monophylétique. Leurs arguments, repris par Stowe
(1986)
(Note 2)
n’étaient pas anatomiques mais uniquement d’ordre
éthologique :
prédation presque exclusive sur des Hétérocères, du moins chez
les Mastophorinae ;
homologie des fils «en ponts»
(«spanning-threads»)
et
des «bolas», peignés
avec les pattes IV lors du
tissage, tous pourvus
d’une glue adhésive
pour les Lépidoptères ;
régurgitations
malodorantes et peut être répugnatoires ; flexion
particulière des pattes I
et II lors du repos
diurne ;
cocons ovigères juxtaposés et à
couche externe
coriace.
Ces
particularités disparates peuvent bien
être considérées comme des synapomorphies
mais
paraissent
néanmoins plus
accessoires que les caractères
anatomiques internes. La structure
fondamentale de l’appareil séricigène reste typique des Araneidae et
n’est donc pas un argument
formel. En revanche, le
tissu folliculaire
endocrinoïde se
présente comme une caractéristique majeure, commune
aux
trois genres Mastophora, Celaenia, Poecilopachys
et
justifiant,
plus que toute autre, leur appartenance à une même série
évolutive. Suggéré
initialement part les
Robinson (1975) et
repris ensuite par Eberhard (1980)
et
l’un des
précédents auteurs (Robinson,1982),
ce groupement
monophylétique dériverait
d’Orbitèles
typiques, incluerait d’abord Poecilopachys et
l’Araignée
néo-guinéenne Pasilobus, réunirait
ensuite les Mastophoreae à
bolas (Mastophora,
Dicrostichus, Cladomelea) pour
s’étendre enfin
aux Celaenieae
qui n'en produisent pas (Celaenia, Taczanowskia).
Il
est capital de souligner ici que la
sous-famille reste
complètement isolée du genre Kaira (Araneinae) au
sein des Araneidae et
ne peut à priori, descendre
d’un même ancêtre. Bien que des
proies leur
soient communes (Lépidoptères
Hétérocères,
Diptères)
et
pareillement attirées par
des odeurs allomonales («mimétisme chimique
agressif"),
trop de caractères
anatomiques et comportementaux s’opposent
à un rapprochement des Mastophorinae
et de Kaira.
Comme le rappelle Stowe
(1986), ce dernier taxon
est
caractérisé
chez le mâle par une apophyse
bulbaire terminale
que ne possèdent pas les
précédents ;
les cocons ovigères
sont superposés, dépourvus de
couche externe coriace et tissés en utilisant
différemment les pattes IV
; il n’existe pas de
régurgitations
malodorantes ; les pattes I et
II ne
sont pas
fléchies au repos.
De plus, les glandes
gnathocoxales ou maxillaires présentent un
dimorphisme sexuel,
inexistant chez Mastophora où
il va
de pair
avec
l’absence de
coiffe. La toile
est rudimentaire mais d’un type et d’une origine
absolument uniques chez les Araignées : les
zigzags déposés
sur sa charpente en trapèze
sont
le produit
des glandes flagelliformes et
surtout, par leur enduit, de glandes
agrégées extraordinairement
modifiées : les
glandes botryoïdes (Lopez,1986). Les
proies
sont
attirées chez Kaira
par
ce même enduit «odorant»,
donc
par la
toile et les glandes agrégées
alors qu’elles sont orientées vers le corps même de l’Araignée
par son tissu endocrinoïde et l’activation
tégumentaire dans le cas des Mastophorinae.
Note 1 :
Laboratoire souterrain du CNRS,
Moulis
09200 : fixation des parties étudiées au
glutaraldéhyde à 2,5% dans un tampon au cacodylate,
post-fixation au tétroxyde d’osmium à 0,2% dans le
même
tampon ; inclusion dans le Spurr ; coupes fines au microtome
Reichert OM U2 contrastées par l’acétate d’uranyle
et le citrate de plomb ; examen de ces coupes au microscope
Sopelem sous 50 KV.Travail
effectué avec Lysiane
Juberthie-Jupeau, directeur de recherches au CNRS, et ses
collaboratrices.
Note
2
: Les
Arachnologistes
US avaient presque tous négligé l’anatomie interne des
Araignées pour leurs études systématiques,
biologiques
et phylogéniques lorsque Lopez a réalisé ses
travaux. Leurs résultats semblent
avoir
“choqué” certains d’entre eux, au point d’interrompre
brutalement une collaboration qui s’annonçait
fructueuse avec
M.K.Stowe. Bien qu'associé aux publications, ce dernier a occulté
la découverte de la glande
botryoïde (Lopez,1985a) dans son travail de
1986, privant
ainsi son article (et ceux qui ont suivi) de la seule explication
logique du comportement de
Kaira. 8 ans plus tard, son
maître H.W. Levi (1993) "ignorait"
toujours l'anatomie interne du
même genre bien qu'elle eut été capitale pour sa
mise au point.
BIBLIOGRAPHIE
Bonaric,
J.C.,
1980.- Thèse Doct.Sci. Univ. Sci. Techn, Montpellier, 282 pp.
Clyne,
D., 1973.-
Aust.ent.Mag.,1(3), April 1973, p.23-30.
Eberhard,
W.G.,1977.- Science,
198, p.1173-1175.
Eberhard,W.G.,
1980.- Psyche,
87
(n°3-4), p.143-169.
Eberhard,W.G.,1981.-
Bull.Br.Arachnol. Soc.,5, p.175-176.
Gertsch,
W.J., 1947.- Natural
Hist.,April 1947, 56 (n°4), p.152-158, 189.
Hutchinson, C.E.,
1903.- Sci.Am., 89
(10), p.172.
Kovoor,J.&
A.Lopez,1982.- Rev.Arachnol.,
4,
p.1-21.
Legendre,R.,
1959.-
Ann.Sci.nat.,Zool. 1 (12), p.339-473.
Legendre,R.,1981.- Rev.Arachnol.,3
(3), p.133-138.
Levi,
H.W., 2003. –
Bull.Mus.Comp. Zool., 157 n° 5, p.309-382.
Lopez,A.,1981 (avec
R.Legendre).- Observations histologiques complémentaires chezl’Araignée
Liphistiomorphe Heptathela kimurai Kishida, 1923
(Liphistiidae).
Atti.Soc.Tosc.Sci ;Nat., Mem., ser.B, 88, suppl.p. 34-44.
Lopez,A.,1985a
(avec M.K.Stowe &
J.C.Bonaric).- Publ.Scient.accel., Univ.R.Descartes, n°8, Mars 1985,
p.1-9.
Lopez,A.1985b (avec M.K.Stowe).- Bull.Soc.Et.Sci.nat.Béziers, N.S., X, Vol.51, 1984-1985, p.16-23.
Lopez,A.,1986 (avec
L.Juberthie-Jupeau & M.K.Stowe).- Mém.Soc.roy.belge Ent.,33,
p.119-128.
Lopez,A.,1998.- Bull.Soc.Et.Sci.nat.Béziers, N.S., XVII, Vol.58, 1997-1998, p.9-21.
Mac
Keown,
K.C.,1952.- Australian Spiders. Angus & Robertson, Sydney.
Millot,J.,1926.-
Supplt.Bull.biol.France Belgique, vol.8, 238
pp.
Millot,J.,1930.-
Arch.Anat.microsc ;, 26 (1), p ;43-81.
Millot,J.,1931.-
Zeitsch.Morph.Okol.Tiere, 21 (3/4), p.740-764.
Millot,J.1968.-
Ordre des Aranéides in Traité de Zoologie, P.P.Grassé édit., Masson, p.
589-743.
Peters, H.M.,1955.-
Z.Naturforsch.,103, p.404.
Robinson,M.H.
& B.Robinson,1975.-Zool.J.Linn.Soc.,56 (4),p.301-314.
Robinson,
M.H. 1982.- Monogr.Biol.,42,
p.557-581.
Simon,E.,1892.-
Histoire naturelle des Araignées. Vol. I, Librairie Encyclopédique de
Roret, Paris.
Steinbrecht,
R.A. 1964.- Z.Zellforsch.Mikroskop.Anat., 64, p.227-261.
Stowe,
M.K., 1986.- in Shear W.A.,ed. : Spiders. Web,
Behavior
and Evolution, Stanford Univ.Press, California, p.101-131.
Streble,H.,1966.-
Zool.Jb.Physiol.,
72,p.157-234.