ARAIGNEES A
ALLOMONES
|
|
Couleurs
conventionnelles :
En noir et italiques, termes anatomiques ; en violet,,
noms
génériques et spécifiques ; en vert, noms de
familles et sous-familles ; en
orange,, parties
les plus importantes
et résumés ; en bleu, liens divers.
|
|
Abréviations
conventionnelles :
M.E.B. : (photographie en)
microscopie
électronique à balayage
M.E.T. :
(photographie en) microscopie
électronique à transmission
C.H. : coupe histologique
(microscopie photonique)
|
|
|
| Fig.1 - Toile géométrique
orbiculaire d' Aranéine. Vue totale.
L'Araignée s'est dissimulée (Hérault) |
Fig.2 - Orbe de Micrathena clypeata installée sur le moyeu. Vue partielle de la toile (Sautero-La Levée, Guyane) |
| C, fils de charpente - M, moyeu fermé (Fig.1) et ouvert (Fig.2) - S, spirale captrice visqueuse traversant les secteurs que délimitent des radii (flèches) (© A.Lopez , d'après deux diapositives) | |
Appelée «fil de capture
gluant» par Peters
(1987), la spirale est la
pièce maîtresse du piège. Elle tire son origine de
4 glandes agrégées
dont le
corps sécrète par son épithélium
un liquide
glycoproteinique visqueux,
faiblement acide, et de deux glandes
flagelliformes élaborant un
produit non
gluant, d’aspect élastique. Ces deux catégories de
glandes s’associent en
unités fonctionnelles
qui disparaissent chez les mâles
adultes (Sekiguchi,
1952 ; Peters,1955). Les
canaux excréteurs des
agrégées ont une
structure
particulière (Kovoor
& Zylberberg,1979) et modifient la
composition finale
de la glu, ainsi que son hydratation. Ils aboutissent aux
filières
postéro-latérales pour s’y terminer par des fusules
étroitement rapprochées en
« triades »
(Kovoor & Lopez,1982). La
sécrétion des
flagelliformes est
extrudée en fibres axiales que recouvre la
glu des agrégées.
Les Araneidae
«spécialistes» de
Lépidoptères et de Diptères mâles ont
abandonné totalement l’orbe conventionnelle et sa spirale
captrice visqueuse.
Ils constituent le seul groupe d’Araignées devenues
réllement monophages ou
presque au cours de leur évolution, un phénomène
remarquable et peu connu
encore des Zoologistes.
Ces
Araignées «vedettes»
appartiennent essentiellement à
trois groupes systématiques : les Celaenieae, les
Mastophoreae
(sous-famille des Mastophorinae)
et le genre Kaira, bien
distinct des
deux précédents, d’ailleurs inclus dans une autre
lignée d’ Araneidae avec
les
genres Metepeira, Aculepeira et Amazonepeira.
Un
quatrième groupe d’ Araneidae,
la tribu des Cyrtarachnineae
(Simon,1892), serait
apparenté aux Mastophorinae (Eberhard,1980 ;
Robinson,1982)
mais n’en a
pas moins conservé le tissage d’une toile
géométrique à orbe modfiée.
Des points communs
indiscutables rapprochent
curieusement les Celaeniae,
les Mastophoreae, les Cyrtarachnineae et Kaira
sans
préjuger des «surprises» que réserve
l’anatomie interne et qui contribueront
ensuite à bien séparer le genre des trois tribus, compte
tenu par ailleurs de
la structure des cocons et de l’éthologie.
► Le corps de
presque toutes
ces Araignées porte de curieuses protubérances
abdominales et prosomatiques,
mousses ou épineuses (Fig.3).
 |
| Fig.3 - Mastophora cornigera, femelle, vue frontale |
| Deux cornes bifurquées sur le
prosoma et protubérances sur l'abdomen. D'après Internet. |
L'histologie montre
que chez Mastophora
celles du céphalothorax,
en forme de "cornes"
bifurquées,
"hébergent" des diverticules intestinaux du thoracentéron
avec leurs
guanocytes ou cellules
à guanine (Fig.4).
|
|
|
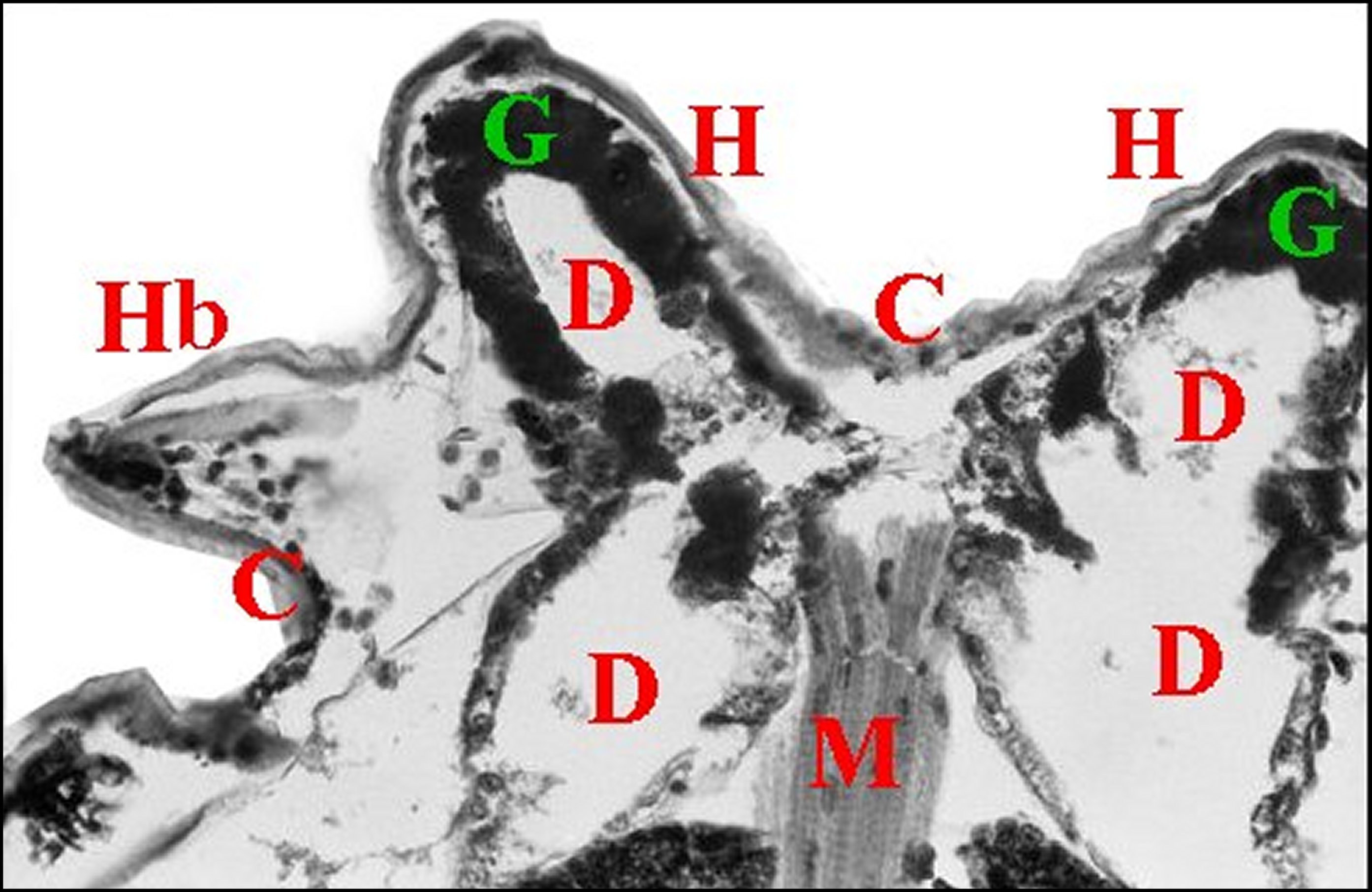
Fig.4 - Mastophora cornigera,
femelle immature : coupe histologique transversale du
prosoma
passant par les cornes (H, Hb). C, cuticule - D, diverticules
intestinaux - G, guanocytes -; M, muscles(© A.Lopez
C.H.) |
►Leurs pattes I et
II sont garnies d’épines
ventrales chez Celaenia (Fig.5), de
longs poils
sétacés, et dans le cas particulier des mâles et
immatures de Mastophora,
d’une rangée prolatérale de soies spéciales
acérées facilitant la
préhension directe des proies (Gertsch,1955 ;
Eberhard,1980)(Fig.6).
 |
 |
| Fig.5 - Celaenia excavata
femelle sous ses cocons |
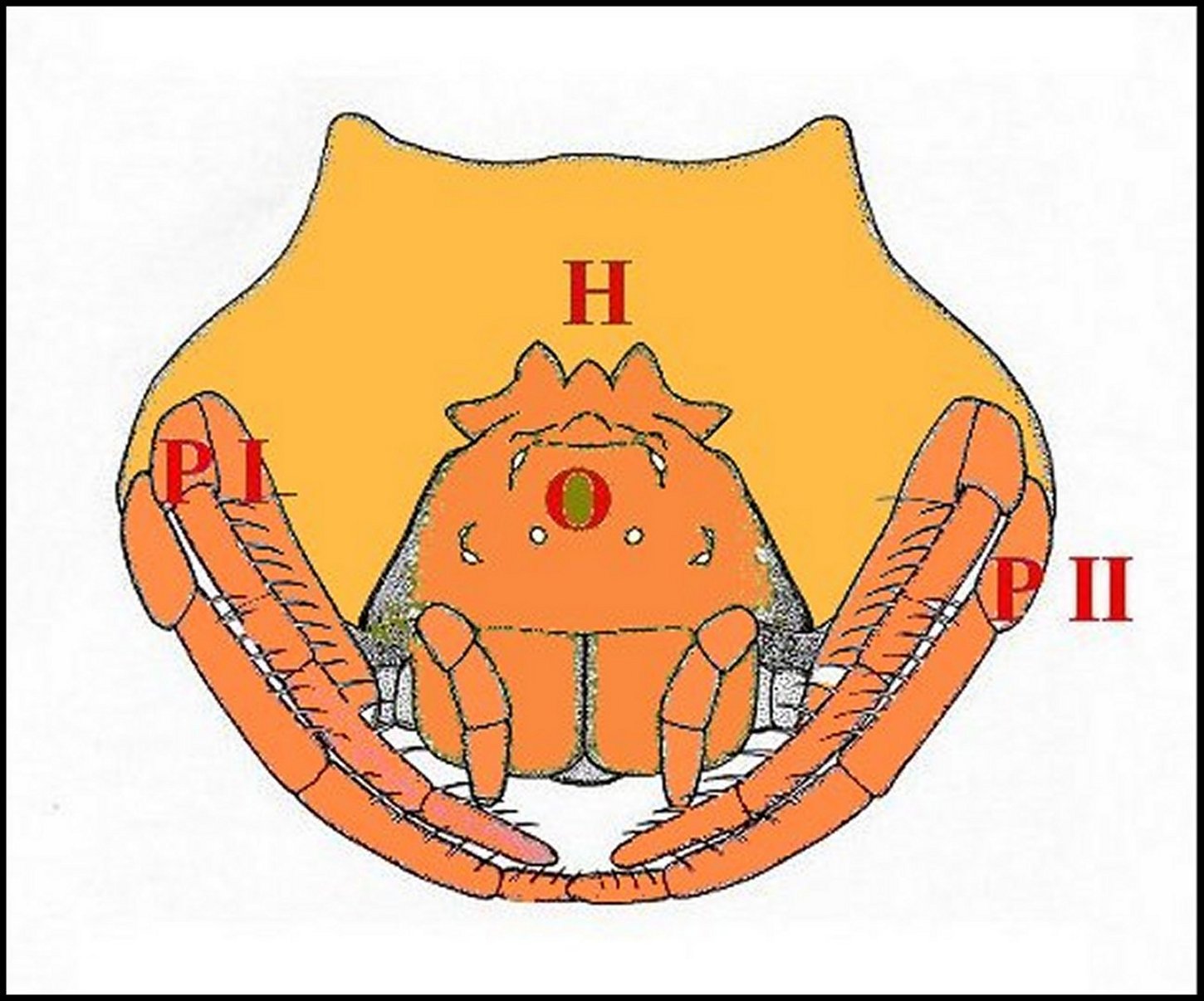
Fig.6 - . Mastophora immature, vue de face : céphalothorax avec ses cornes, pattes I et II, abdomen avec ses bosses à l'arrière plan . |
| Pattes hérissées
d'épines ventrales (fémurs), étalées dans
l'attente d'une proie (©B.Lopez,
Brisbane) |
H, cornes - 0, région oculaire. Les pattes sont garnies de soies (©A.Lopez, dessin.) |
►Le dimorphisme
sexuel
volumétrique est souvent considérable :
mâles nains
ou «pygmées» de Mastophora
(Gertsch,
1955) (Fig.7,8)
et Celaenia
sans atteindre toutefois
l’inégalité de taille
extravagante observée chez d’autres Araneidae telles que les Argiope, Cyrtophora et les Nephila
(Néphiles) (Fig.9).
►Elles se camouflent de jour dans
la végétation grace à leurs formes et couleurs
cryptiques (Fig.10), parfois vives (Poecilopachys)(Fig.12)
mais pouvant aussi les faire ressembler à des…fientes d’oiseau
(« bird
dropping spiders ») lorsque le blanc et le
noir prédominent comme chez Caelenia (Fig.11) !
 |
 |
 |
|
Fig.10 - Mastophora
cornigera
Femelle, vue dorsale.
Flèche
: cornes
|
Fig. 11 - Celaenia excavata
Femelle,
vue dorsale (Col.A.Lopez)
|
Fig.12-
Poecilopachys australasia
Femelle, vue
dorsale (Col.A.Lopez)
|
►De
même,
elles tendent à dissimuler leurs gros cocons ovigères,
fusiformes ou globuleux,
sessiles ou pédiculisés, parfois ornementés,
qu’elles tissent parmi les plantes
(Fig. ).
► Leur activité
prédatrice est strictement
nocturne, ce qui en rend l’observation aléatoire et très
difficile.
►Les femelles adultes capturent des
Hétérocères
nocturnes, exclusivement dans
le cas des Mastophorinae
et de Kaira, peut
être aussi d’autres
proies chez les Cyrtarachneae.
Ces Papillons, du moins en Amérique, se rattachent
à 7 familles d’
Hétérocères : Noctuidae, Geometridae,
Tineidae, Plutellidae,
Tortricidae, Gelechiidae et Pyralidae ( in
Yeargan, 1994).
►En revanche, les jeunes immatures,
les mâles subadultes
des Mastophorinae, de Kaira
et
même adultes de certaines
Mastophora ne capturent
pas des Lépidoptères mais des Diptères
Nématocères, presque toujours du sexe
mâle, dont la taille est plus en rapport avec la leur :
Cecidomyidae dans
le cas de Kaira
alba et
surtout Psychodidae dans celui des Mastophora
et de Celaenia
(Hickman, 1970 ; Forster & Forster,1973 ;
Lauder in
Eberhard,1980 ; Yeargan &
Quate,1996). Une relation
spécio-spécifique
existerait d’ailleurs chez trois Mastophora
différentes (Yeargan
& Quate,1996), chacun de ces
taxons tendant
à ne capturer qu’une ou deux espèces de Psychoda.
►Il existe en gros trois modes de capture
excluant totalement
l’orbe classique.
-Utilisation d’une toile rudimentaire
en trapèze pourvue
d’un «zigzag» singulier chez Kaira, du moins
la femelle de K.
alba.
-Elaboration
de fils pendants
garnis d’un
ou plusieurs globules trés visqueux et manipulés
avec
une patte ambulatoire
, ainsi que l'a bien décrit Gertsch (1947) comme
les «bolas» qu'utilisent les gauchos
sud-américains pour entraver leur bétail.
-Absence
de tout dispositif
soyeux de
capture chez les Celaenieae, ce que corrobore leur appareil
séricigène réduit.
-Pour leur part les
Cyrtarachneae
(in Cyrtarachninae) construisent une
toile géométrique élaborée
sub- horizontale, rappelant les
orbes conventionnelles d’autres Araneidae par sa charpente, son moyeu
et ses
radii. Elle est toutefois dépourvue de spirale gluante captrice
que
remplacent des fils tendus «en ponts» trés
lâches dans les
secteurs(« spanning-threads »), garnis en leur
centre d’un matériel
extrèmement visqueux, pendant après rupture comme des
«bolas» et engluant
surtout des Lépidoptères
Hétérocères. Ces derniers pourraient être
attirés par
des éléments de la toile et (ou) l’araignée qui
l’a construite.
►Sous
toute réserve pour
les Cyrtarachneae, dont
l’éventail des proies est encore mal connu, les Insectes
capturés sont presque invariablement des mâles. Les
Araignées émettent pour les
«appeler» des substances volatiles odorantes,
diffusées seulement pendant la
chasse (Gemeno & al., 2000)
et qui simulent les
phéromones sexuelles
de leurs femelles. Ces sémiochimiques font donc partie des
allomones.puisqu’ils
sont utilisés pour la communication entre espèces
différentes et provoquent une
réponse adaptative favorable pour
l’ «émetteur-prédateur», léthale
pour la «proie-receveur »
►Il existe
diverses preuves de la «duperie chimique»
(«agressive chemical mimicry» : Eberhard,1977 ;
Yeargan,1988)
pratiquée par les Mastophorines
et
Kaira.
Les
mâles des Insectes,
Diptères et
surtout Lépidoptères, se dirigent vers l’ Araignée
qui les «appelle» en volant
contre le vent et en contournant les obstacles visuels
interposés, ce qui
prouve bien que l’attraction est olfactive.
Les
Papillons appartiennent aux
seules
espèces d’Hétérocères qui soient actives
sexuellement dans le secteur lors de
la scotophase où l’ Araignée les capture (2 seulement sur
71 : Eberhard,
1977). Certains d’entre eux déploieraient même leurs genitalia
lorsqu’ils approchent du prédateur, confirmant ainsi que
l’odeur
attractive ne peut être que sexuelle, et cela, d’autant que la
reproduction
est leur unique activité imaginale. Qui plus est, les
Lépidoptères mâles
qu’attire Kaira alba
pourraient ébaucher une pseudo-copulation avec la toile
collabée de cette Araneidée (obs.
de Stowe). Ceci ne serait pas sans rappeler la tentative d'
"accouplement" des mâles de certains Insectes
Hyménoptères (Scoliidae) avec le labelle d' Ophrys
(Orchidaceae) pour polliniser cette dernière !
Il
est par ailleurs
démontré que certains
composés chimiques phéromonaux appartenant aux
groupes HCE (hydrocarbures
ou époxydes) et surtout AAA (aldéhydes aliphatiques,
alcools, acétates (in
Yeargan, 1994 ; Haynes & al., 1996) sont
produits à la
fois par
certains Lépidoptères nocturnes et par des Mastophoreae
qui les capturent (Stowe &
al.,1987 ;
Yeargan,1994). En fait ces dernières
élaboreraient
plutôt des mélanges
phéromonaux («pheromone blends» : Stowe &
al.,1987) dont la
composition peut varier dans le temps et expliquerait la capture
successive
d’espèces différentes au cours d’une même nuit.
►Le
lieu de diffusion phéromonale a
été
recherché expérimentalement par les auteurs
nord-américains qui l’ont attribué
chez les Mastophorinae, non au matériel des bolas, du
moins lorsqu’ils
existent, mais au corps même de l’
Araignée (Eberhard,1977) et,
chez Kaira, à la seule toile (Stowe, com.pers.).
Il n’en restait pas moins que
la source
glandulaire des attractifs allomonaux, problème majeur,
demeurait inconnue
jusqu’en 1985. A partir de cette même année, l'auteur a
donc
effectué une série de
recherches histologiques pour tenter de la mettre en évidence
(Lopez,1985a ;
Lopez,1985b ;
Lopez,1986 ;
Lopez,1998 ;
Lopez,1999)
Elles se
sont
avérées d’emblée
fructueuses dans le genre Kaira
(Lopez,1985a), y montrant une
correlation aussi
spectaculaire que logique entre l’anatomie des glandes
à soie agrégées, converties en glandes botryoïdes pourvues de
coussinets
à cellules géantes (Fig.7,8) et le
comportement prédateur
de Kaira alba (Lopez,1985b ; Lopez,1986).
Beaucoup
plus tard seulement,
ces recherches ont
orienté A.Lopez
(1998)
vers une autre
conception du phénomène chez les Mastophorinae dont le tissu endocrinoïde folliculaire
abdominal,
situé près des glandes à soie (Fig.9,10), y est sans nul doute impliqué. On peut aujourd’hui
même ( dès 2007) l’étendre aux
Cyrtarachneae sur la base
de documents histologiques antérieurs
à 1997.
Eberhard,W.G.,
1980.- Psyche, 87 (n°3-4),
p.143-169.
Forster,R.R.
& L.Forster, 1973.- New Zealand
Spiders, Collins ed.,
London.
Gemeno,C.,
Yeargan,K.V. &
K.F.Haynes, 2000.- Chemical Ecology, 26,p.1235-1243.
Gertsch, W.J.,
1947.- Natural Hist.,April 1947, 56
(n°4),
p.152-158, 189.
Gertsch,W.J.,
1955.-
Bull.Amer.mus.nat.Hist.,106 (4), p.221-254.
Haynes,K.F., Yeargan, K.V., Millar,J.G. & B.B.Chastain,1996.- Journ.Chem.Ecol.,22 (n°1),p.75-89.
Hickman,V.V.,1970.- Pap.Proc.Royal Soc.Tasm.,105, p.75-81.
Hutchinson,
C.E., 1903.- Sci.Am., 89 (10),
p.172.
Kovoor,
J. &
L. Zylberberg,1979.-
Zoomorph.,92,p.217-239.
Kovoor,J. & A.Lopez,1982.- Rev.Arachnol., 4, p.1-21.
Lopez,A.,1985a (avec M.K.Stowe et J.C.Bonaric).- Publ.Scient.accel., Univ.R.Descartes, n°8, Mars 1985, p.1-9.
Lopez,A.,1999.-
Alexanor, 21,
fasc.4, p. 195-200.
Peters,
H.M.,1955.- Z.Naturforsch.,103, p.404.
Peters,
H.M.,
1987.- in Nentwig,W.,
ed. : Ecophysiology of Spiders, Springer-Verlag, Berlin, V, p.
187-202.
Robinson,M.H.
&
B.Robinson,1975.-Zool.J.Linn.Soc.,56 (4),p.301-314.
Robinson,
M.H. 1982.- Monogr.Biol.,42,
p.557-581.
Sekiguchi,
K.1952.- Annot.Zool.Japan.,25, p.394-399.
Simon,E.,1892.- Histoire naturelle des Araignées. Vol. I, Librairie Encyclopédique de Roret, Paris.
Stowe,M.K., Tumlinson, J.H. & R.R.Heath,1987.- Science, 236, p.964-967.
Allomones
and Kairomones: Transspecific Chemical Messengers. BioScience, Volume 20, Issue 1, January 1970, p.21
Yeargan,K.V,
1988.- Oecologia (Berlin),74p.524-530.
Yeargan,
K.V.
& L.W.Quate,1996.-
Oecologia, 106 (2), p. 266-271.
Yeargan,
K.V.,1994.- Ann.Rev.Entomol.,39,p.81-99.