 |
 |
 |
| Fig. 1 - Scytodes
thoracica, femelle, vue dorsale |
Fig. 2 - Même femelle, autre
vue dorsale |
Fig. 3 - Autre femelle, transportant
sa ponte. Vue latérale droite |
| Flèche rouge : "gibbosité "
dorsale |
Flèche jaune : "gibbosité" ;
flèche rouge : chélicères |
P, ponte appliquée sur le sternum
par les
palpes. Flèche : "gibbosité" |
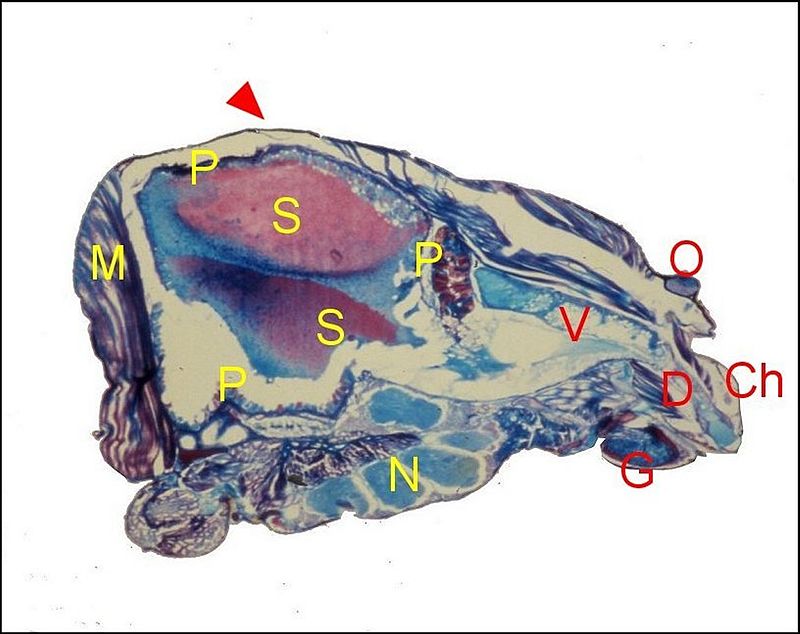
Enorme
et bilobée, elle présente dans les coupes histologiques
une trés large lumière et surtout deux poches, dont la
structure et
la sécrétion ont un aspect différent : une
poche antérieure élaborant
toujours du venin ; une poche postérieure
plus grande,
devenue réellement séricigène et
sécrétant la substance visqueuse que projettent les
chélicères (Fig.4
). Le canal excréteur est
commun aux deux.
 |
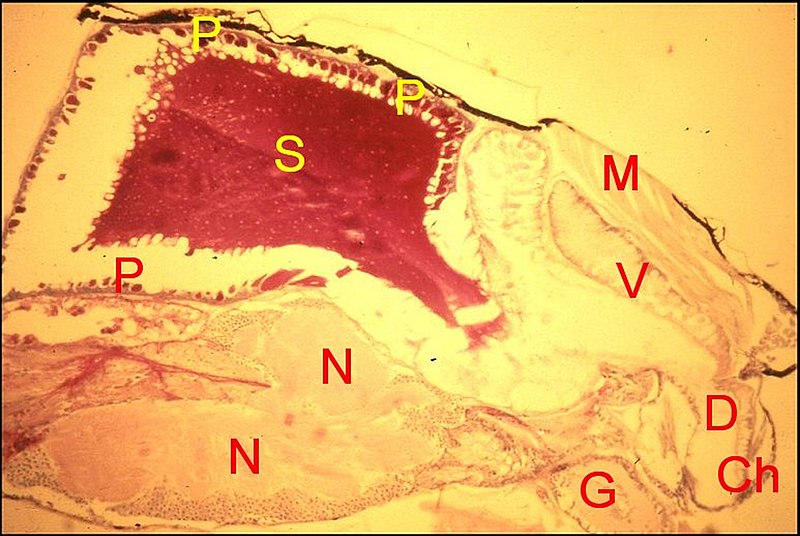 |
| Fig. 4 - Scytodes
thoracica, femelle : coupes histologiques parasagittales du
céphalothorax. Trichrome, à gauche ; A.P.S., à
droite. |
|
| Ch,
chélicère ; D, canal
excréteur; G, gnathocoxe ; M, muscles; N, ganglions nerveux ; O,
oeil ; P, paroi de la
poche séricigène ; S, sa sécrétion ; V,
portion antérieure restée venimeuse. Flèche :
"gibbosité" |
|
Les filières, qui représentent sur le plan phylogénétique des appendices modifiés, sont situées près de l'extrémité de l'abdomen, en avant du tubercule anal. Généralement au nombre de trois paires, antérieure, moyenne et postérieure, chacune d'elles montre respectivement 3,1 et 2 segments.
Leur disposition est des plus variables, parfois presque en un seul plan chez les Hahnia (Fig. ). Leur longueur peut être trés importante, surtout dans le cas des Hersilia dont la paire postérieure semble démesurée (Fig. ).
Les filières sont pourvues d'une musculature bien développée leur assurant une grande mobilité, aussi bien individuelle que coordonnée en travail d'ensemble durant la construction des cocons et des toiles.Une 4eme paire, médiane antérieure et siégeant sur le segment 10, est considérée comme ancestrale et n'a persisté, fonctionnelle, que chez quelques Mesothelae. En revanche, elle a subi une réduction extrême dans le cas de beaucoup d' Araignées écribellates comme les Araneidae (Fig.5), les Linyphiidae, les Theridiidae, les Theridiosomatidae (Fig.6) et les Thomisidae ; elle s'y présente comme une saillie vestigiale, impaire et médiane, appelée colulus, manquant dans d'autres groupes (Fig.7), mais sans raison apparente, car cet organe ne parait pas exercer de fonction .
 |
 |
 |
| Fig. 5-Cyrtophora citricola, femelle :
filières. M.E.B. |
Fig.
6 - Theridiosoma sp., femelle
: filières. M.E.B. |
Fig. 7 - Araignée sp.,
femelle : filières. M.E.B. |
| A,
filières antérieures ; C, colulus ; M, filières
médianes ; P, filières postérieures ; T, tubercule
anal |
||
 |
 |
| Fig. - Hahnia
sp. (Réunion) , femelle : filières, les
postérieures réclinées. M.E.B. |
Fig. -
Hersilia
savignyi , femelle : vue partielle de filière
postérieure. M.E.B. |
| A, filières antérieures ; F, fusules; H, poils ; M, filières médianes ; P, filières postérieures ; T, tubercule anal | |
Les fusules (Fig.8 à 10) ont l'aspect de "canules" effilées recevant chacune un canal excréteur qui la parcourt axialement jusqu'à son extrémité libre et s'y ouvre par un pore. Le "corps" est plus ou moins strié mais non cannelé. L'embase a une forme conique ou cylindrique (Fig. 8 à 10). Elles rappellent beaucoup celles des glandes épigastriques prégonoporales.
 |

|
 |
| Fig. 8 - Cyrtophora citricola : filière postérieure. M.E.B. | Fig.9 - Argyrodes sp.
: Fusules d'une filière médiane. M.E.B. |
Fig.10 - Argyrodes sp. : filière médiane. M.E.B. |
| A, alvéoles de poils ; F, fusules ; H, poils ; P, pore apical de fusule ; S1, segment externe de la filière ; S2, segment interne. Flèche : embase cylindrique ou conique de fusule. | ||
Chez Telema tenella (Telemidae) par exemple, il y a trois paires de filières, décrites brièvement par Fage (1913) mais dont notre étude en M.E.B a fourni une description plus complète (Lopez,1983a)
Les filières antérieures encadrent un large colulus, sont allongées, cylindriques et comportent deux articles. Le terminal est pourvu de 7 fusules coniques, à embase annulaire, réparties sur un plateau oblique : 6 d'entre elles, recevant probablement les canaux des glandes piriformes, y sont groupées deux par deux ; la septième est isolée, plus courte et plus interne.......;
Les Araignées dites cribellates possèdent en outre un appareil fileur additionnel, le cribellum, situé juste en avant des filières classiques.
Plusieurs catégories principales de glandes peuvent être individualisées sur les plans morphologique (Schéma 1) et histologique (Fig.11) : les glandes ampullacées, aciniformes, piriformes, agrégées, tubuliformes et flagelliformes
Chaque catégorie de glande élabore une soie présentant ses propres caractères spécifiques et a ainsi une fonction particulière.
- Les glandes piriformes aboutissent toutes aux filières antérieures (antéro-latérales) et produisent les disques de fixation pour la toile.
- Les glandes ampullacées aboutissent aux filières antérieures et moyennes ; elles produisent les fils de charpente de la toile ainsi que les fils de rappel ("drag lines")
- Les glandes aciniformes aboutissent aux filières moyennes et postérieures ; elles élaborent les soies d'enveloppement des proies, de la toile spermatique (chez le seul mâle) et de la paroi externe du sac ovigère
- Les glandes tubuliformes se terminent également dans les filières moyennes et postérieures ; elles ne produisent que la soie des cocons.
- Les glandes agrégées, considérées comme propres aux Araneidae se retrouvent aussi chez les Tetragnathidae, les Linyphiidae, les Theridiidae et élaborent la glue visqueuse quel qu'en soit le support, y compris dans les "bolas" de Mastophora. Elles sont remarquablement modifiées en glandes botryoïdes dans le genre Kaira.
- Les glandes flagelliformes sécrètent le fil qui supporte la glue et s'associent généralement aux agrégées en "triade fonctionnelle" (Schéma 2)








































